Фізіологія людини - Вільям Ф. Ґанонґ 2002
Фізіологія нервових і м'язових клітин
Збудлива тканина: нервова
Нейротрофні речовини
Трофічні фактори нервів
Виділено і вивчено велику кількість білків, потрібних для життєдіяльності і росту нейронів. Окремі з них - нейротрофіни - утворені м’язами або іншими структурами, що їх іннервують нейрони, інші виділені астроцитами. Ці білки зв’язуються з рецепторами нервових закінчень, інтерналізуються і ретроградно переміщуються до тіла нервової клітини, де сприяють синтезу білків, потрібних для розвитку, росту і життєдіяльності нейронів. Інші нейротрофіни утворюються в нейронах, антероградно транспортуються до нервових закінчень; вони потрібні для нормального функціювання постсинаптичного нейрона.
Рецептори
У табл. 2-4 наведені чотири достеменно визначені нейротрофіни та їхні високоафінні рецептори. Кожен з цих Trk рецепторів димеризується і таким способом ініціює автофосфорилювання в доменах цитоплазматичної тирозинкінази рецепторів. Відомий також додатковий низько-афінний рецептор ФРН - фактор росту нервів; білок з молекулярною масою 75 кДа, його називають p75NTR. Цей рецептор зв’язує всі чотири названі нейротрофіни з однаковим ступенем афінності. Є підстави вважати, що він здатний утворювати гетеродимер з мономером TrkA. Такий димер підвищує афінність і специфічність зв’язування фактора росту нервів. Однак з’ясовано: гомодимери p75NTR, які зв’язують нейротрофіни, можуть самі бути посередниками реакцій, що ведуть до апоптозу - явища, протилежного до звичайного ростостимулювального і трофічного ефекту нейротрофінів.
Таблиця 2-1. Типи нервових волокон у нервах ссавців1
Тип волокна |
Функція |
Діаметр волокна, мкм |
Швидкість передавання, м/с |
Тривалість піка, мс |
Період абсолютної рефрактерності, мс |
А а |
Пропріоцептивна, соматомоторна |
12-20 |
70-120 |
0,4-0,5 |
0,4-1,0 |
ß |
Дотик, тиск |
5-12 |
30-70 |
||
y |
Рухова щодо м’язових веретен |
3-6 |
15-30 |
||
δ |
Біль, холод, дотик |
2-5 |
12-30 |
||
В |
Прегангліонарне автономне волокно |
<3 |
3-15 |
1,2 |
1,2 |
С Задній корінець |
Біль, температура, окремі види механорецепції, рефлекторні відповіді |
0,4-1,2 |
0,5-2 |
2 |
2 |
Симпатичне |
Постгангліонарне симпатичне волокно |
0,3-1,3 |
0,7-2,3 |
2 |
2 |
1 А- і В-волокна мієлінові, С-волокна безмієлінові.
Фактор росту нервів
Першим дослідженим нейротрофіном був фактор росту нервів (ФРН) білкового типу, потрібний для росту та функціювання симпатичних і деяких чутливих нейронів. Цей фактор є в багатьох тваринних видів, у тім числі і в людини: міститься в різних тканинах. Особливо високу його концентрацію виявлено в підщелепній залозі самців миші. Після кастрації самців концентрація ФРН зменшується до рівня, властивого самкам. Фактор побудований з двох а-, двох ß- і двох у-субодиниць. У ß-субодиницях, молекулярна маса кожної з яких 13 200, є уся ростостимулювальна активність, а-субодиниці виявляють трипсиноподібну дію, a у-субодиниці - це серинпротеази. Функція цих протеаз нез’ясована. Структура ß-одиниці ФРН нагадує структуру інсуліну. ФРН проникає в нейрони з позачерепних органів, що їх іннервують ці нейрони, і переміщується ретроградно від нервових закінчень до тіл нервових клітин. Крім того, ФРН міститься в головному мозку й, очевидно, необхідний для росту і життєдіяльності холінергічних нейронів основи переднього мозку та смугастого тіла. Ін’єкція антитіл до ФРН новонародженим тваринам спричинює майже повну деструкцію симпатичних вузлів, зумовлюючи імуносимпатектомію. Можна вважати, що під впливом ФРН явища апоптозу в нейронах менше виражені.
Таблиця 2-2. Цифрова система, яку інколи використовують для класифікації волокон чутливих нейронів
Номер |
Початок волокна |
Тип волокна |
Іа |
М’язове веретено, кільцево-спіральне закінчення |
Аа |
Ів |
Сухожильний орган Ґольджі |
Аа |
II |
М’язове веретено, гроноподібне розширення; рецептори дотику, тиску |
Aß |
III |
Рецептори болю, холоду і деякі рецептори дотику |
Аδ |
IV |
Рецептори болю, температури й інші |
С-волокно заднього корінця |
Інші нейротрофіни
Продукований мозком нейротрофний фактор (BDNF - від англ. brain-derived neurotropic factor), нейротрофін-3 (NT-3), NT-4/5 і ФРН забезпечують різні сторони життєдіяльності нейронів, запобігаючи апоптозу, хоча їхні функції перекриваються. Створення дефіциту NT-3 шляхом нокауту гена спричинює помітну втрату механорецепторів у шкірі (див. Розділ 7) навіть у гетерозиготних організмах. BDNF діє швидко і може фактично деполяризувати нейрони. Дефіцит BDNF у мишей спричинює втрату периферійних чутливих нейронів і глибокі дегенеративні зміни в вестибулярних вузлах, а також пригнічення LTP (див. Розділ 4). Описаний також фактор NT-6.
Інші фактори росту нейронів
Клітини Шванна й астроцити виділяють циліарний нейротрофний фактор (CNTF - від англ. ciliary neurotropic factor), який сприяє виживанню ушкоджених і зародкових нейронів спинного мозку; тому його можна застосовувати для лікування хвороб, у разі яких відбуваються дегенеративні зміни в нейронах. Нейротрофний фактор, продукований лінією гліальних клітин (GDNF - від англ. glial cell line-derived neurotpopic factor), активізує дофамінергічні нейрони середнього мозку in vitro. У тварин з нокаутом GDNF дофамінергічні нейрони виглядають нормальними. Однак у них нема нирок і порушений розвиток ентеричної нервової системи (див. Розділ 26). Іншим фактором, що сприяє росту нейронів, є інгібувальний фактор лейкемії (LIF - від англ. leukemia inhibitory factor). Крім того, нейрони, як і інші клітини, реагують на інсуліноподібний фактор росту І (ІФР-І), різні форми трансформувального фактора росту (ТФР), фібробластний фактор росту (FGF - від англ. fibroblast growth factor) і фактор росту тромбоциту (PDGF - від англ. platelet-derived growth factor). Отже, регулювання росту нейронів є складним процесом.
Таблиця 2-3. Порівняльна чутливість нервових волокон ссавців А, В і С типів до різних інгібіторів провідності
Чутливість до |
Найбільше чутливі |
Середньо-чутливі |
Найменше чутливі |
Гіпоксія |
В |
А |
С |
Стискання |
А |
В |
С |
Анестетики місцевої дії |
С |
В |
А |
Таблиця 2-4. Нейротрофіни
Нейротрофін |
Рецептор |
Фактор росту нервів |
TrkA |
Нейротрофний фактор, продукований мозком |
TrkB |
Нейротрофін 3 |
TrkC, меншою мірою TrkA і TrkB |
Нейротрофін 4/5 |
TrkB |
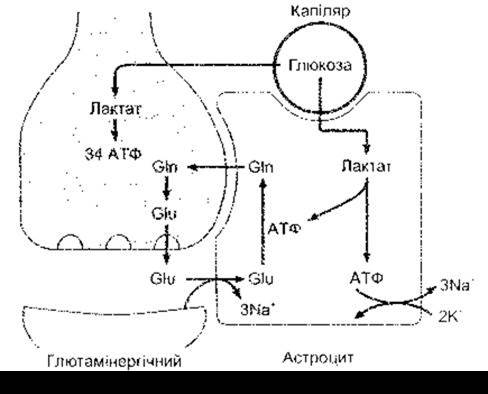
Рис. 2-15. Глютамат-глютаміновий цикл між глютамінергічними нейронами й астроцитами. Глютамат (Glu), що виділився в синаптичну щілину, зв’язує Nа+-залежний глютаматний транспортер і в астроциті перетворює на глютамін (Gln). Глюкоза з капілярів проникає в астроцити і нейрони. В астроцитах вона метаболізується до лактату з утворенням двох молекул АТФ. Одна з них забезпечує перетворення глютамату на глютамін, а іншу використовує Nа+-К+-АТФ-аза для переміщення трьох йонів Na+ з клітини в обмін на два йони К+. У нейронах глюкоза метаболізується в циклі лимонної кислоти з утворенням 34 молекул АТФ.
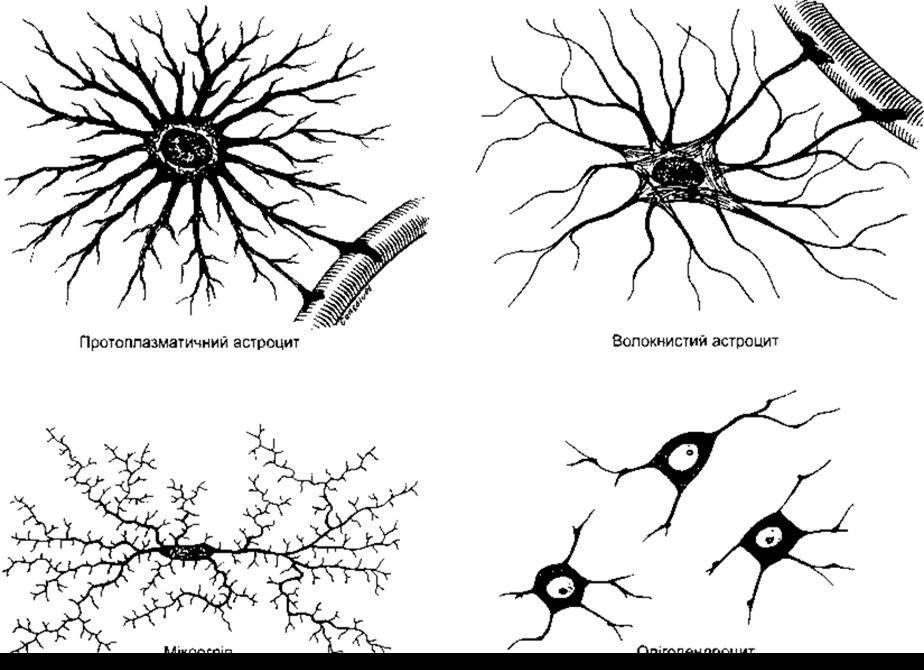
Рис. 2-14. Гліальні клітини головного мозку (відтворено за дозволом з Junqueira L, Саmеіrо J, Kelley R Basic Histology, 9th ed. McGraw-Hill, 1998).