Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції травної системи
Травлення та всмоктування
Вуглеводи
Травлення
Головними харчовими вуглеводами є полісахариди, дисахариди та моносахариди. Крохмаль (полімер глюкози) та інші похідні - це єдині полісахариди, що перетравлюються до будь-якого ступеня в травному тракті людини. В глікогені більшість молекул глюкози мають лінійні довгі ланцюги (молекули глюкози, об’єднані 1,4-а глікозидними зв’язками), хоча інколи бувають і розгалужені ланцюги (сформовані завдяки 1,6-а-зв’язкам; див. рис. 17-12). Амілопектин, який становить 80-90% харчового крохмалю, є нерозгалуженим полісахаридом, тоді як амілоза - лінійний полісахарид, у якому мономери сполучені прямим ланцюгом, має винятково 1,4-а-зв’язки. Глікоген міститься в тварин, тоді як амілоза й амілопектин - рослинного походження. Дисахариди лактоза (молочний цукор) і сахароза (головний компонент харчового цукру, найпоширеніший у природі) теж перетравлюються до моносахаридів фруктози і глюкози.
У ротовій порожнині крохмаль взаємодіє з а-амілазою слини. Оптимальне pH для дії цього ензиму - 6,7, отже, його активність гальмує кисле pH шлункового соку, коли їжа потрапляє до шлунка. В тонкій кишці обидві а-амілази (як слинна, так і панкреатична) діють на поглинуті полісахариди. Ці а-амілази гідролізують 1,4-а-зв’язки, однак зберігають і 1,6-а-зв’язки, кінцеві 1,4-а- і наступні до місця розгалуження 1,4-а. Кінцевим продуктом травлення а- амілазою є олігосахариди: дисахарид мальтоза, трисахарид мальтотріоза, деякі легкі великі полімери з глюкозою в 1,4-а-зв’язку, а-декстрини - полімери глюкози із розгалуженою будовою, що містять у нормі понад вісім молекул глюкози, основний тип зв’язку - 1,6-а глікозидний (рис. 25-1). Олігосахариди, відповідальні за майбутнє травлення похідних крохмалю, містяться на зовнішній ділянці щіточкової облямівки - мембрані з мікроворсинками тонкої кишки (рис. 25-2). Деякі з цих ензимів мають понад один субстрат. Ензим а-декстриназа, який також відомий як ізомальтаза, головно відповідальний за гідроліз 1,6-а- зв’язків.
Таблиця 25-1. Головні травні ензими. Відповідні проензими наведені в круглих дужках
Джерело |
Ензим |
Активатор |
Субстрат |
Каталітична функція або продукт |
Слинні залози |
Слинна а-амілаза |
Cl- |
Крохмаль |
Гідроліз 1,4-а-зв’язків, продукування а-лімітованих декстринів, мальтотріози і мальтози |
Язикові залози |
Лінгвальна ліпаза |
Тригліцериди |
Жирні кислоти плюс 1,2-діацилгліцерол |
|
Шлунок |
Пепсини (пепсиногени) |
HCl |
Білки і поліпептиди |
Роз’єднання пептидних зв’язків, суміжних до ароматичних амінокислот |
Шлункова ліпаза |
Тригліцериди |
Жирні кислоти і гліцерол |
||
Підшлункова залоза, Екзокринний відділ |
Трипсин |
Ентеропептидаза |
Білки і поліпептиди |
Роз’єднання пептидних зв’язків на карбоксильній стороні основних амінокислот (аргінін або лізин) |
Хімотрипсин (хімотрипсиноген) |
Трипсин |
Білки і поліпептиди |
Роз’єднання пептидних зв’язків на карбоксильній стороні ароматичних амінокислот |
|
Еластаза (проеластаза) |
Трипсин |
Еластин, деякі інші протеїни |
Роз’єднання зв’язків на карбоксильній стороні аліфатичних амінокислот |
|
Карбоксипептидаза А (прокарбоксипептидаза А) |
Трипсин |
Білки і поліпептиди |
Роз’єднання карбоксильних кінцевих амінокислот, що мають ароматичні або розгалужені аліфатичні сторони ланцюгів |
|
Карбоксипептидаза В (прокарбоксипептидаза В) |
Трипсин |
Білки і поліпептиди |
Роз’єднання карбоксильних кінцевих амінокислот, що мають основну сторону ланцюгів |
|
Коліпаза (проколіпаза) |
Трипсин |
Крапельки жирів |
Сприяє реалізуванню дії активних сайтів панкреатичної ліпази |
|
Панкреатична ліпаза |
Тригліцериди |
Моногліцериди і жирні кислоти |
||
Холестеролестер гідролаза |
Ефіри холестеролу |
Холестерол |
||
Панкреатична а-амілаза |
Cl- |
Крохмаль |
Так само, як слинна а-амілаза |
|
Рибонуклеаза |
РHК |
Нуклеотиди |
||
Дезоксирибонуклеаза |
ДНК |
Нуклеотиди |
||
Фосфоліпаза А2 (профосфоліпаза А2) |
Трипсин |
Фосфоліпіди |
Жирні кислоти, лізофосфоліпіди |
|
Слизова оболонка кишки |
Ентеропептидаза |
Трипсиноген |
Трипсин |
|
Амінопептидаза |
Поліпептиди |
Відщеплення амінокислот з амінокінців пептидів |
||
Карбоксипептидаза |
Поліпептиди |
Відщеплення амінокислот з карбоксидних кінців пептидів |
||
Ендопептидази |
Поліпептиди |
Роз’єднання між залишками всередині пептиду |
||
Дипептидази |
Дипептиди |
Дві амінокислоти |
||
Мальтаза |
Мальтоза, мальтотріоза, а-декстрини |
Глюкоза |
||
Лактаза |
Лактоза |
Галактоза і глюкоза |
||
Сахараза1 |
Сахароза, також мальтотріоза і мальтоза |
Фруктоза і глюкоза |
||
а-Декстриназа |
а-Декстрини, мальтоза, мальтотріоза |
Глюкоза |
||
Трегалаза Нуклеаза і споріднені ензими |
Трегалоза Нуклеїнові амінокислоти |
Глюкоза Пентози і пуринові та піримідинові основи |
||
Цитоплазма слизових клітин |
Різноманітні пептидази |
Ди-, три- і тетрапептидази |
Амінокислоти |
1 Сахараза і а-декстриназа є від’єднаними субодиницями від окремого білка.
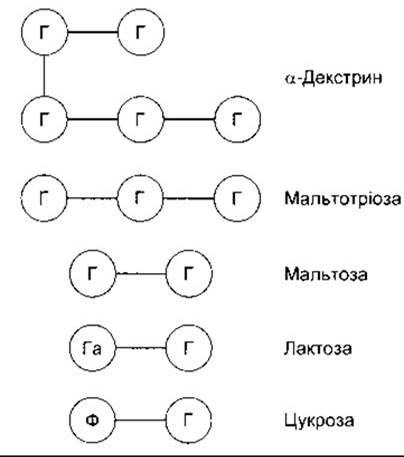
Рис. 25-1. Головні кінцеві продукти травлення вуглеводів у просвіті кишки. Кожне коло зображує молекулу гексози; Г - глюкоза; Ф - фруктоза; Га - галактоза.
Разом з мальтазою і сахаразою ензими також розчеплюють мальтотріозу та мальтозу. Сахараза та а-декстриназа, первісно синтезовані як окремий глікопротеїновий ланцюг, долучаються до мембранного краю щіточкової облямівки. Потім панкреатичні протеази його гідролізують до субодиниць сахарази та ізомальтази, які нековалентно реасоціюють до поверхні кишки.
Сахараза також гідролізує сахарозу в молекули глюкози і фруктози. Додатково в щіточковій облямівці є два дисахариди: лактаза, яка гідролізує лактозу до глюкози та галактози, і трегалаза, яка гідролізує тригалозу - 1,1 -а-ланцюговий димер глюкози - до двох молекул глюкози.
Дефіцит одного або більше олігосахаридів щіточкової облямівки може спричинювати діарею, здуття та метеоризм після вживання солодощів. Діарея зумовлена збільшенням кількості осмотично активних молекул олігосахаридів, що залишаються в просвіті кишки, сприяючи збільшенню об’єму інтерстинального вмісту. В ободовій кишці бактерії розщеплюють деякі олігосахариди, у подальшому збільшуючи кількість осмотично активних частинок. Процеси гниття та бродіння в дистальній частині тонкої і товстої кишки, відповідно, продукують гази (СО2 і Н2) із залишків дисахаридів.
Інтерес до лактази зумовлений тим, що у більшості ссавців і в багатьох расах людей активність кишкової лактази висока під час народження, а потім знижується в дитячому та дорослому віці. Низький рівень лактази асоціюється з несприйняттям молока (несприйняття лактози). Більшість європейців та їхні американські потомки зберігають активність інтестинальної лактази в зрілі роки. Випадки дефіциту лактази для північних та західних європейців становлять 15%, однак для афроамериканців та американських індіанців, орієнтальців та середземноморської популяції - 70-100%. Проблему несприйняття молока можна частково вирішити за допомогою комерційних препаратів лактази, проте це дорого. Особи з непереносимістю ліпше сприймають йогурти, ніж молоко завдяки безпосередньому вмісту власної бактеріальної лактази.
Абсорбція
Гексози і пентози швидко всмоктуються через стінку тонкої кишки (табл. 25-2). По суті, виведення всіх гектоз відбувається вздовж термінальної частини клубової кишки. Молекули цукрів проходять через клітини слизової і потрапляють у кровоносні капіляри, а потім - до портальної вени.

Рис. 25-2. Специфічні субстрати для ензимів, що беруть участь у травленні вуглеводів, та гексози як кінцеві продукти травлення. Числа - відсотковий вміст кожного розщепленого субстрату відповідним ензимом. Зверніть увагу, що трегалаза, лактаза і цукраза - це єдині ензими, що відповідають за розщеплення трегалози, лактози, цукрози, однак а-декстрини, мальтотріоза і мальтоза - субстрати для декількох ензимів (відтворено за дозволом з Johnson LR [editor]: Essential Medical Physiology, Raven, 1992)
Транспортування більшості гексоз унікальне і відбувається за участю Na+, що міститься в кишковому просвіті. Висока концентрація Na+ у поверхні слизової полегшує, а низька - пригнічує надходження цукрів до епітеліальних клітин. Це трапляється тому, що глюкоза і Na+ беруть участь у котранспортуванні, або симпорті, тобто натрійзалежному транспортуванні глюкози (SGLT - від англ. sodium-dependent glucose transporter). Члени цієї родини транспортерів SGLT 1 і SGLT 2, подібно до транспортерів глюкози, відповідають за полегшену дифузію (див. Розділ 19), пронизують клітинну мембрану 12 разів і розміщують свої -СООН і -NН2-кінці на цитоплазматичній стороні мембрани. Крім того, вони негомологічні до серії GLUT-транспортерів. SGLT 1 і SGLT 2 також відповідають за транспортування глюкози з ниркових канальців (див. Розділ 38).
Якщо внутрішньоклітинна концентрація Na+ стає низькою в ентероцитах, як і в інших клітинах, то Na+ рухається за концентраційним градієнтом. Глюкоза надходить разом з Na+ і вивільнюється у клітину (рис. 25-3). Йони Na+ переходять у бічні міжклітинні простори, а глюкоза за допомогою GLUT 2 - до інтерстиційного простору, а звідти - до капілярів.
Отже, транспортування глюкози є прикладом вторинного активного транспортування (див. Розділ 1). Енергетичне забезпечення транспортування глюкози відбувається безпосередньо за допомогою активного транспортування Na+ з клітини. Це підтримує концентраційний градієнт уздовж люмінального краю клітини: більше Na+ і, відповідно, більше глюкози може увійти до неї. Наслідком успадкованої вади Nа+/глюкози котранспортера є розвиток синдрому малабсорбції глюкози/галактози, що спричинює важку діарею, яка часто є фатальною, якщо глюкозу і галактозу не вилучити з раціону харчування. Вживання глюкози та її полімерів утримує Na+ і зумовлює прояви діареї, що описані нижче.
Транспортування галактози також відбувається за допомогою глюкози. Фруктоза утилізується за іншим механізмом. Її реабсорбція не залежить від Na+ або транспортування глюкози чи галактози; це відбувається завдяки полегшеній дифузії: з просвіту кишки до ентероцитів за допомогою GLUT 5 і в зворотному напрямі - за допомогою GLUT 2. Деяка кількість фруктози перетворюється в глюкозу в слизових клітинах. Пентози всмоктуються шляхом простої дифузії.
Інсулін незначно впливає на транспортування цукрів у кишці. З цього погляду інтестинальна абсорбція у кишці подібна до реабсорбції глюкози в проксимальних звивистих канальцях у нирках (див. Розділ 38), не потребує фосфорилювання і обидва процеси простежуються у разі цукрового діабету (гальмує лікарський засіб флоризин). Максимальний рівень всмоктування глюкози в кишці становить 120 г/год.