Фізіологія людини - Вільям Ф. Ґанонґ 2002
Дихання
Транспортування газів між легенями і тканинами
Транспортування оксиду вуглецю
Буферні системи
Для того, щоб зрозуміти процеси буферування в організмі з моменту утворення СО2 із вугільної кислоти у крові, потрібно з’ясувати, як відбувається транспортування СО2. Буферні системи описані у Розділі 1 і детально розглянуті у Розділі 39.
Рух оксиду вуглецю в крові
Розчинність СО2 у крові в 20 разів більша, ніж розчинність О2, тому вважають, що за однакових парціальних тисків СО2 є більше, ніж О2 у простому розчині. У випадку дифузії в еритроцити СО2 швидко гідратується до Н2СО3, оскільки наявна карбоангідраза. Внаслідок дисоціації Н2СО3 утворюються Н+ і НСО3. Йон Н+ буферує головно гемоглобін, а НСО3 потрапляє у плазму. Частина СО2 в еритроцитах реагує з аміногрупами білків, насамперед з гемоглобіном, і утворює карбаміносполуки.
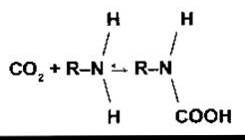
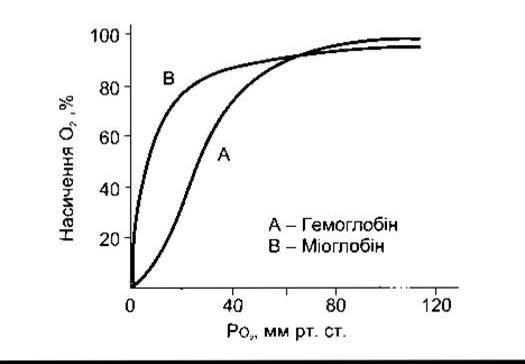
Рис. 35-5. Криві дисоціації гемоглобіну і міоглобіну.
Як тільки деоксигенований гемоглобін зв’яже більше Н+, ніж це робить оксигемоглобін, розпочинається швидке утворення карбаміносполук, зв’язуючи О2 з гемоглобіном і зменшуючи його спорідненість з СО2 (ефект Галдана). Венозна кров переносить більше СО2, аніж артеріальна, і поглинання СО2 полегшується у тканинах, а СО2 вивільнюється у легенях. Понад 11% СО2 долучається у капілярну кров великого кола кровообігу і переноситься до легень як карбаміно-СО2
У плазмі СО2 реагує з білками плазми, утворюючи невелику кількість карбаміносполук, незначна частина СО2 гідратує. Реакція гідратації відбувається повільно без карбоангідрази.
Переміщення хлору
Унаслідок збільшення вмісту НСО3 в еритроцитах порівняно з плазмою під час проходження крові через капіляри понад 70% цього НСО3 переходить у плазму. Рух надлишку НСО3, що виходить з еритроцитів в обмін на Сl- (рис. 35-6), контрольований білком смужки 3, головним мембранним білком. Цей обмін називають переміщенням хлору, оскільки завдяки йому вміст Сl- в еритроцитах венозної крові значно більший, ніж в артеріальній. Переміщення хлору відбувається швидко і повністю закінчується за 1 с.
Зверніть увагу, що кожна молекула СО2, яка надходить в еритроцити, збільшує їх на одну осмотично активну частинку: або НСО3, або Сl (див. рис. 35-6). Внаслідок цього еритроцити набирають воду і збільшуються у розмірі. З цього погляду, а також беручи до уваги, що невелика кількість рідини в артеріальній крові повертається через лімфатичні судини швидше, ніж через вени, гематокрит венозної крові у нормі на 3% більший, ніж артеріальної. У легенях Сl виводиться з клітин і вони зморщуються.
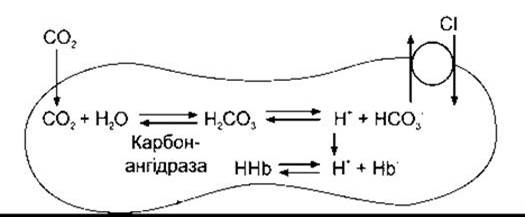
Рис. 35-6. Зміни, що відбуваються в еритроцитах у разі додавання СО2 у кров. Зверніть увагу, що кожна молекула СО2, яка входить у еритроцити, - це додатковий йон НСО3- або Cl- у клітину.
Підсумок
З огляду на різноманітність функцій СO2 у плазмі та еритроцитах їх систематизовано у табл. 35-2. З рис. 35-7 видно, що збільшення ємності крові переносити СO2 визначене різницею між лініями, що позначають розчинений СO2 і загальний СO2 у кривій дисоціації для СO2.
У кожному декалітрі артеріальної крові є близько 49 мл СO2 (див. табл. 35-1): 2,6 мл розчиненого, 2,6 мл утворює карбаміносполуки і 43,8 мл - у НСO3-. У тканинах на кожний декалітр крові припадає 3,7 мл СO2, 0,4 мл залишається у розчині, 0,8 мл утворює карбаміносполуки і 2,5 мл - CHO3. Показник pH крові з 7,4 зменшується до 7,36. У легенях відбувається протилежний процес, і 3,7 мл СO2 вивільняється через альвеоли. У такий спосіб у спокої 200 мл CO2 за 1 хв і значно більше під час навантаження транспортується від тканин до легень і виводиться. Це заслуговує на увагу, бо така кількість СО2 еквівалентна надолужити 12 500 мекв Н+ за 24 год.
Таблиця 35-2. СO2 у крові

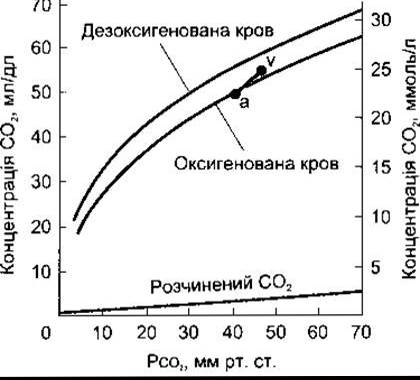
Рис. 35-7. Крива дисоціації С02. Артеріальна (а) і венозна точки (v) відображають загальний вміст СO2 в артеріальній і венозній крові людини у нормі в стані спокою (модифіковано та відтворено за дозволом з Schmidt RF, Thews G [editors]: Human Physiology. Springer, 1983).