Фізіологія людини - Вільям Ф. Ґанонґ 2002
Утворення і виділення сечі
Функція нирок і сечовипускання
Окиснювання сечі та екскреція бікарбонату
Секреція Н+
Клітини проксимальних і дистальних відділів ниркових канальців, подібно до клітин шлункових залоз, виконують секрецію йонів Н+ (див. Розділ 26). Ацидифікація відбувається також у збірних трубочках. Головним процесом, за допомогою якого секретуються H+ у проксимальних канальцях, є Na+-H+ обмін (рис. 38-20). Це приклад вторинного активного транспортування; Na+-K+-ATO-aзa витісняє йони Na+ з клітин у міжклітинну рідину, що призводить до зменшення внутрішньоклітинної концентрації Na+. Завдяки цьому Na+ починає надходити всередину клітин із канальцевої рідини; у цьому разі відбувається витіснення одного йона Н+. Йони H+ утворюються внаслідок внутрішньоклітинної дисоціації Н2СO3, а НСO3-, що виникає в цьому випадку, шляхом дифузії потрапляє в міжклітинну рідину. Отже, в обмін на секрецію кожного йона H+ у міжклітинну рідину надходить один йон Na+ і один йон НСO3-.

Рис. 38-20. Секреція кислот клітинами проксимального відділу ниркового канальця; Н+ переходить у просвіт канальця за допомогою антипорта в обмін на йон Na+. Активне транспортування за допомогою Nа+-К+-АТФ-ази позначено стрілками у колі. Штриховими лініями позначено дифузію. Порівняйте з рис. 26-10.
Каталізатором реакції утворення Н2СO3 є ензим карбангідраза. Медикаментозні лікарські препарати, які інгібують його, водночас пригнічують секрецію Н+ у проксимальних відділах ниркових канальців і всі пов’язані з цим біохімічні реакції.
Уважають, що в секреції Н+ у проксимальній частині канальця беруть участь також інші нагнітальні системи, однак їхнє існування однозначно не доведено. В будь-якому випадку, вони відіграють незначну роль порівняно з Na+- Н+-обмінним механізмом. У дистальній частині канальця та збірних трубочках простежується цілком протилежна ситуація, адже там секреція Н+ практично не залежить від концентрації Na+ в канальцевій рідині. У цих ділянках канальця більшу частину Н+ секретує АТФ-залежна протонна помпа. Альдостерон діє на цю помпу, збільшуючи секрецію H+ у дистальній частині канальця. Вставні клітини (І-клітини) в цій ділянці ниркового канальця секретують H+ і, подібно до парієтальних клітин слизової шлунка, містять значну кількість карбангідрази та численні тубуловезикулярні структури. В літературі є дані про те, що в цих пухирцях і у внутрішній мембрані епітеліальних клітин канальців міститься так звана Н+-транслокувальна АТФ-аза, яка, власне, і продукує H+. У разі ацидозу кількість протонних помп збільшується внаслідок вставлення цих тубуловезикул в повернений до просвіту канальця бік клітинної мембрани. Деяку кількість H+ секретує Н+-К+-АТФ-аза. У базолатеральній мембрані І-клітин міститься так званий Band 3 - аніоно-обмінний білок, який, імовірно, відіграє роль С -НСО3 -обмінника в процесі перенесення НСО3- у міжклітинну рідину
Метаболізм Н+ у сечі
Інтенсивність секреції кислот залежить від процесів, описаних нижче. Для нирок кожного біологічного виду характерний максимальний градієнт концентрації Н+, за якого транспортні механізми ще здатні виконувати активне транспортування. У людини цей градієнт відповідає pH сечі близько 4,5, тобто концентрації йонів Н+, яка в 1000 разів перевищує їхню концентрацію в плазмі крові. Отже, pH 4,5 - це граничне значення pH сечі. В нормі саме таким є рН рідини у збірних трубочках. Якби не було буферних систем, які «зв’язують» Н+ у сечі, то граничне значення pH досягалось би дуже швидко, і секреція Н+ припинялась би. Однак є три важливі хімічні реакції, за допомогою яких надлишок вільних йонів Н+ легко виходить з канальцевої рідини, що дає змогу виконувати подальшу секрецію кислот (рис. 38-21). Це реакції з НСО3 із утворенням СО2 і Н2О, із НРО42 з утворенням Н2РО4, та з NH3 із утворенням NH4+.
Реакції з буферними системами
Механізм роботи буферних систем розглянутий у Розділах 1 та 39. Значення рК’ бікарбонатної системи становить 6,1, фосфатної - 6,8, аміачної - 9,0. Концентрація НСО3 у плазмі, і, як наслідок, у клубочковому фільтраті в юрмі становить 24 мекв/л, тоді як концентрація фосфату лише 1,5 мекв/л. Отже, у проксимальній частині канальця більша частина H+ реагує із НСО3 з утворенням Н2СО3 див. рис. 38-21). Відповідно, Н2СО3 розпадається на СО2 Н2О. Щіточкова облямівка клітин проксимального (але не дистального) канальця містить карбангідразу, це полегшує утворення СО2 і Н2О в канальцевій рідині. Водночас СО2, який легко дифундує через усі біологічні мембрани, надходить у канальцеві клітини, де він поповнює пул СО2, використовуваний для синтезу Н2СО3. Оскільки переважна більшість Н+ виходить із канальця, то pH канальцевої рідній помітно не змінюється. Власне цей процес є механізмом, а допомогою якого відбувається реабсорбція НСО3-: на кожен моль НСО3, що виходить з канальцевої рідини, шляхом дифузії 1 моль НСО3 переходить з канальцевих клітин кров, хоч це не є той самий моль, який перейшов від анальцевої рідини.
Виділений H+ взаємодіє з двоосновним фосфатом НРО42 ), унаслідок чого утворюється одноосновний фосфат (Н2РО4). Цей процес найбільше виражений у дисальній частині канальця і у збірних трубочках, бо власне унаслідок реабсорбції води накопичується найбільша кількість йонів фосфату, які не були реабсорбовані в просимальній частині канальця. Реакція з NH3 відбувається к у дистальній, так і в проксимальній частинах канальця: он Н+ з’єднується й з іншими буферними аніонами, та значно менше.
Кожен йон Н+, що реагує з буферними системами, є внеском до так званої титраційної кислотності сечі, яку визначають як кількість лугу, який треба додати до сечі, щоб її pH становив 7,4, що відповідає pH клубочкового фільтрату. Проте очевидно, що титраційна кислотність відображає лише частину Н+, яка була секретована, оскільки в цьому разі не врахована Н2СО3, що перетворилась на Н2О і СО2. Крім цього, рК' амонійної системи становить 9,0, а вона титрується тільки від pH сечі до 7,4, отже, її внесок до титраційної кислотності сечі є дуже незначним.
Секреція аміаку
Унаслідок хімічних реакцій у клітинах ниркових каналь- ців утворюються NH4+ І НСО3-. Йон NH4+ перебуває у стані рівноваги із системою NH3 + Н+ усередині клітин. Оскільки рК' цієї реакції дорівнює 9,0, то співвідношення між NH3 і NH4+ при pH 7,0 становить 1:100 (рис. 38-22). Крім того, NH3 є жиророзчинною сполукою і тому за концентраційними градієнтами легко переходить через клітинні мембрани у міжклітинну рідину і канальцевий фільтрат. У сечі він реагує з Н+, утворюючи NH4+, який залишається в сечі.
Головною реакцією, унаслідок якої в клітинах утворюється NH4+, Є перетворення глутаміну у глутамат. Каталізатор цієї реакції - ензим глутаміназа, наявний у значній кількості в клітинах ниркових канальців (див. рис. 38-22). Глутамінова дегідрогеназа каталізує перетворення глутамату в а-кетоглутарат з утворенням додаткової кількості NH4+. З подальшим перетворенням а-кетоглутарату споживається 2 Н+, унаслідок чого вивільнюється 2 НСО3-.

Рис. 38-21. Метаболізм Н+, секретованого у канальцеву рідину в обмін на Na+. Угорі: реабсорбція бікарбонатів за допомогою СО2. У центрі: утворення одноосновного фосфату. Унизу: утворення аміаку. Зверніть увагу, що на кожен секретований йон Н+ виділяється один Na+ і один НСО3-; А- - аніон.
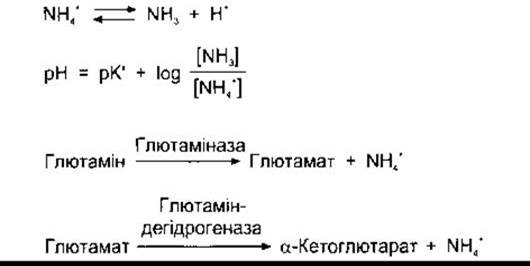
Рис. 38-22. Головні реакції утворення аміаку в нирках. Див. також Розділ 17.
У разі хронічного ацидозу екскреція NH4+ посилюється. Це простежується за будь-якого значення pH, оскільки більша кількість NH3 надходить у канальцеву рідину. Це явище отримало назву адаптації секреції NH3, його причину ще остаточно не з’ясовано. Адаптація секреції NH3 призводить до посилення секреції Н+.
Процес, завдяки якому NH3 секретує у сечу і в подальшому перетворюється на NH4+, унаслідок чого підтримується концентраційний градієнт NH3, називають нейонною дифузією (див. Розділ 1). Секреція саліцилатів та деяких інших медикаментів, які є слабкими основами чи слабкими кислотами, відбувається також шляхом нейонної дифузії. Швидкість їхнього екскретування залежить від pH сечі.
Зміни pH у різних відділах нефрона
У проксимальному канальці простежується незначне зменшення pH, та, як зазначено вище, екскреція Н+ не має значного впливу на pH канальцевої рідини з причини утворення СО2 і Н2О з Н2СО3. На противагу цьому, клітини дистальної частини канальця значно менше здатні до секреції Н+, однак секреція саме на цьому відрізку найбільше впливає на pH сечі.
Фактори, що впливають на секрецію Н+
Секреція кислот з сечею змінюється під впливом PCО2 всередині клітини, концентрації К+, рівня карбангідрази і концентрації гормонів кори надниркових залоз. Якщо РСО2 високий (респіраторний ацидоз), то більше Н2СО3 доступно для зв’язування гідроксильних йонів і секреція кислот посилюється, тоді як зі зниженням РСО2 простежуються протилежні явища. Нестача К+ зумовлює посилення екскреції кислоти, очевидно, тому, що втрата К+ призводить до внутрішньоклітинного ацидозу навіть у випадку, коли pH плазми крові збільшене. Навпаки, надлишок К+ всередині клітини пригнічує секрецію кислот. Під час інгібування карбоангідрази секреція кислот також зменшена, оскільки зменшується утворення Н2СО3. Альдостерон, а також інші гормони кори надниркових залоз, які посилюють канальцеву реабсорбцію Na+, паралельно підвищують секрецію Н+ і К+.
Екскреція бікарбонатів
Хоча в процесі реабсорбції НСО3 не відбувається транспортування цього йона в канальцеві клітини, реабсорбція НСО3 пропорційна до його фільтрації. Нема чітко визначеного Тм, проте реабсорбція НСО3 послаблюється за допомогою невідомого механізму у випадках, коли збільшується об’єм ПКР (рис. 38-23). Якщо концентрація НСО3 у плазмі крові мала, то НСО3 реабсорбується повністю. Однак якщо концентрація НСО3 перевищує 26-28 мекв/л (нирковий поріг для НСО3-), то НСО3 з’являється в сечі, і сеча стає більш лужною. Навпаки, коли концентрація НСО3 в плазмі крові менше 26 мекв/л (значення, за якого всі секретовані йони Н+ використовуються для реабсорбції НСО3-), то вивільняється більше Н+, які можуть зв’язуватися з аніонами інших буферних систем. Отже, чим менша концентрація НСО3 у плазмі крові, тим кислішою стає реакція сечі і тим більший вміст у ній NH4+.
Фізіологічне значення змін pH сечі
Залежно від тісно взаємнопов’язаних процесів секреції Н+, продукування NH4+ і екскреції НСО3 pH сечі у людини коливається в широких межах - від 4,5 до 8,0. Екскреція сечі, pH якої значно відрізняється від pH рідких середовищ організму, має важливе значення для електролітного та кислотно-лужного балансу організму, що детально розглянуто в Розділі 39. Кислоти зв’язуються в плазмі крові і клітинах, рівняння хімічної реакції виглядає так: НА + NaHCО3 —> NaA + Н2СО3. Відповідно, Н2СО3 розпадається на СО2 і Н2О; СО2 всмоктується в кров і виходить під час видихання, як NaA з’являється в клубочковому фільтраті. Та кількість Na+, яка заміщується в сечі на Н+, надходить назад в організм. На кожен йон Н+, що піддався екскреції з фосфатом або у вигляді NH+4, організм отримує один йон НСО3-, завдяки чому поповнюються запаси цього важливого буферного аніона. Натомість, коли до рідких середовищ організму надходить луг, то йони ОН зв’язуються з утворенням додаткової кількості НСО3-. Якщо ж концентрація НСО3 в плазмі крові перевищує 28 мекв/л, то сеча стає лужною і надлишок НСО3 виходить з нею. Оскільки ж максимальна швидкість секреції Н+ в канальцях напряму залежить від РСО2 в артеріальній крові, то на реабсорбцію НСО3 значно впливає РСО2 Детальніше про ці взаємовідношення описано у Розділі 39.
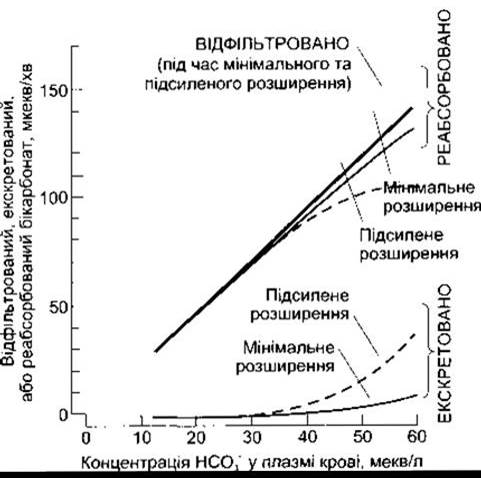
Рис. 38-23. Вплив об’єму ПКР на фільтрацію, реабсорбцію та екскрецію НСО3- у дослідах на щурах. У людей екскреція НСО3- відбувається подібно. В нормі концентрація НСО3- у плазмі крові становить близько 24 мекв/л (відтворено за дозволом з Valtin Н: Renal Function, 2nd ed. Little, Brown and Co., 1983)