Фізіологія людини - Вільям Ф. Ґанонґ 2002
Утворення і виділення сечі
Функція нирок і сечовипускання
Екскреція води
У нормі клубочки за добу фільтрують 180 л рідини, а середній добовий діурез становить у середньому 1 л. Така ж кількість розчинених речовин може за добу виводитись в об’ємі сечі 500 мл з концентрацією 1400 мосм/кг чи в об’ємі сечі 23,3 л з концентрацією 30 мосм/кг (табл. 38-7).
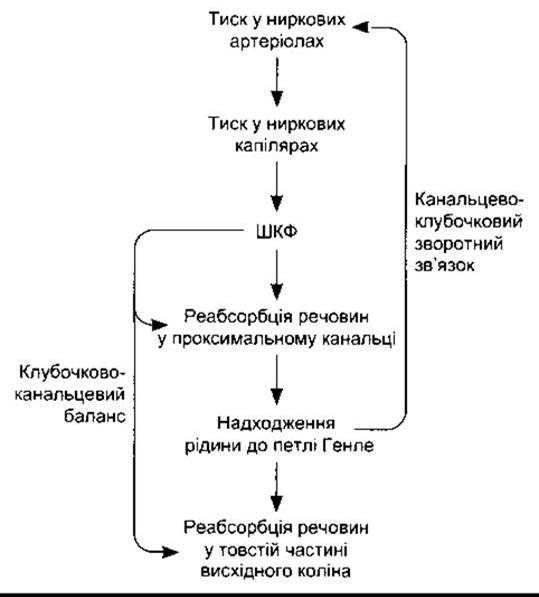
Рис. 38-14. Механізми клубочково-канальцевого балансу та канальцево-клубочкового зворотного зв’язку.
Таблиця 38-7. Зміни метаболізму води, спричинені дією вазопресину в людей. У всіх випадках осмотичне навантаження екскретованої рідини становить 700 мосм/день
ШКФ, мл/хв |
Відсоток реабсорбції води |
Діурез, л/добу |
Концентрація сечі, мосм/кг Н2О |
Втрата або надлишок рідини, л/добу |
|
Ізотонічна плазмі сеча |
125 |
98,7 |
2,4 |
290 |
|
Вазопресин (максимальний антидіурез) |
125 |
99,7 |
0,5 |
1400 |
+1,9 |
Відсутність дії вазопресину (нецукровий діабет) |
125 |
87,1 |
23,3 |
30 |
-20,9 |
Наведені цифри свідчать про два важливі факти: по-перше, реабсорбується принаймні 87% води, навіть якщо добовий діурез становить 23 л; по-друге, реабсорбція води може коливатись у широких межах без впливу на екскрецію розчинених у ній речовин. Отже, якщо сеча концентрована, то вода затримується в організмі, якщо ж сеча низької концентрації, то вода виходить. Обидва ці факти мають важливе значення для раціонального використання води і підтримання сталості осмотичного тиску рідких середовищ організму. Водний баланс регулює здебільшого вазопресин, який впливає на збірні трубочки.
Проксимальний відділ канальця
У проксимальному відділі канальця відбувається транспортування багатьох речовин, проте, за даними мікропункцій, осмотичний тиск рідини вздовж усього проксимального відділу канальця дорівнює осмотичному тиску плазми крові (див. рис. 38-9). Отже, в проксимальному відділі канальця вода реабсорбується пасивним шляхом за осмотичними градієнтами, що утворилися внаслідок активного транспортування розчинених речовин, і таким способом, підтримується ізотонічність рідини. Рух води полегшує наявність водних каналів в апікальних мембранах епітеліальних клітин проксимального відділу канальця; HgCl2 зворотно інгібує водні канали. Ці канали в проксимальному відділі канальця складаються з білка аквапорину-1. Сьогодні виділено чотири додаткові представники цієї родини білків. Вони забезпечують рух води шляхом простої дифузії. Аквапорин-2, який на 42% гомологічний аквапорину-1, локалізований у клітинах збірних трубочок. Він є трансмітером відповіді цих клітин надію вазопресину (див. нижче і Розділ 14). Аквапорин-3 локалізований у базолатеральній частині мембран збірних трубочок і бере участь у транспортуванні сечовини, гліцерину і води. Аквапорин-4 виявляють у мозку, а аквапорин-5 - у слинних і слізних залозах, а також у дихальній системі.
Хоча вазопресин не впливає на аквапорин-1, цей білок відіграє важливу роль у збереженні води. У разі його нокауту в дослідах на мишах проникність проксимального відділу канальця для води знижується на 80%, тварини втрачають до 35% маси тіла; у разі збезводнення осмотичний тиск плазми у них підвищується до 500 мосм/кг Н2O, незважаючи на те, що аквапорини-2, -3, і -4 є інтактними.
Концентрація інуліну (який не реабсорбується) в рідині у кінці проксимального відділу канальця в 2,5-3,3 раза більша, ніж у плазмі крові. З цього випливає, що під час проходження через проксимальний відділ канальця реабсорбується 60-70% води і розчинених у ній речовин (рис. 38-15).
Петля Генле
Як зазначено вище, петлі юкстамедулярних нефронів значно заглиблюються в мозкові піраміди, перш ніж впадають у дистальні звивисті канальці в кірковому шарі, і всі збірні трубочки знову проходять скрізь ниркові піраміди, впадаючи через верхівки пірамід у ниркову миску. В міжклітинній рідині пірамід простежується поступове збільшення осмоляльності: осмоляльність у ділянці верхівок пірамід становить близько 1200 мосм/кг Н2O, що майже в чотири рази перевищує осмоляльність плазми. Низхідне коліно петлі Генле проникне для води, а висхідне - ні (табл. 38-8). Йони Na+, К+ і Сl реабсорбуються з товстого сегмента висхідного коліна шляхом котранспортування (див. нижче). Отже, якщо вода переходить у гіпертонічну міжклітинну рідину, то рідина у низхідному коліні петлі Генле також стає гіпертонічною. У висхідному коліні вона більше розведена і, якщо вона досягає верхівки, то вже має осмотичний тиск, нижчий від плазми внаслідок руху Na+ і Сl за межі просвіту канальця. Під час проходження через петлю Генле реабсорбується ще близько 15% води, тому в дистальний відділ канальця надходить лише 20% від попередньої кількості води, і концентрація інуліну тут уже в п’ять разів переважає його концентрацію в плазмі крові.
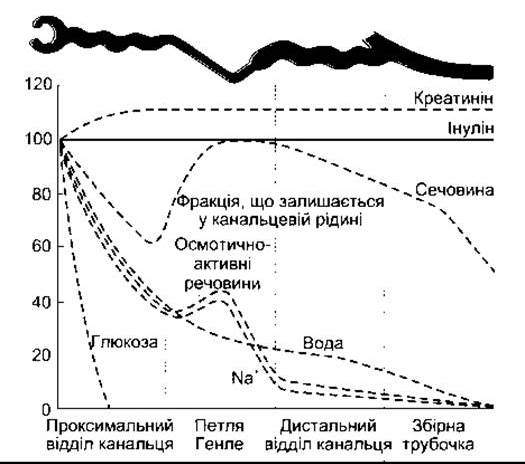
Рис. 38-15. Зміни відсоткової частки відфільтрованої кількості речовин, що залишаються у канальцевій рідині уздовж нефрона (відтворено за дозволом з Sullivan LP, Grantham JJ: Physiology of the Kidney, 2nd ed. Lea&Febiger, 1982).
Таблиця 38-8. Проникність і транспортування різних сегментів нефрону1
Проникність |
Активне транспортування Na+ |
|||
Н2O |
сечовина |
NaCl |
||
Петля Генле |
||||
Тонке низхідне коліно |
4+ |
+ |
± |
0 |
Тонке висхідне коліно |
0 |
+ |
4+ |
0 |
Товсте висхідне коліно |
0 |
± |
± |
4+ |
Дистальний звивистий каналець Збірна трубочка |
± |
± |
± |
3+ |
Кіркова частина |
3+* |
0 |
± |
2+ |
Зовнішня мозкова частина |
3+* |
0 |
± |
1 + |
Внутрішня мозкова частина |
3+* |
3+ |
± |
1 + |
1 Результати отримані з досліджень нирок кроля і людини. Дані, позначені *, отримані під впливом вазопресину; +1 - за відсутності вазопресину (модифіковано та відтворено за дозволом з Kokko JP: Renal concentrating .and diluting mechanisms. Hosp Pract [Feb] 1979; 110:14).
Функція транспортного механізму у товстому висхідному коліні петлі Генле залежить від носія, який з просвіту канальця переносить всередину клітин один йон Na+, один йон К+ і два йони Сl (рис. 38-16). Крім того, Na+-K+-ATO-аза активно виводить Na+ з клітин у міжклітинний простір; у цій ділянці ниркового канальця вміст цього ензиму є найвищим; К+ переміщається назад у просвіт канальця шляхом простої дифузії. Один йон Сl пасивно дифундує в міжклітинний простір, а ще один переміщається разом з К+.
Білок, що виконує транспортування Na+-K+-2Cl у товстій частині висхідного коліна петлі Генле, має 12 трансмембранних доменів. їхні амінові і карбоксильні кінці повернуті досередини клітини. Цей білок належить до родини транспортерів, які містяться в багатьох інших органах, у тім числі слинних залозах, шлунково-кишковому тракті і дихальних шляхах.
Дистальний відділ канальця
Дистальний відділ канальця, особливо його початкова частина, є, фактично, продовженням товстого сегмента висхідного коліна петлі Генле. Він порівняно непроникний до води, і подальша реабсорбція розчинених речовин зі збереженням розчинника призводить до ще більшого розведення канальцевої рідини. Ця ділянка канальця дає всього 5% у загальну реабсорбцію води.
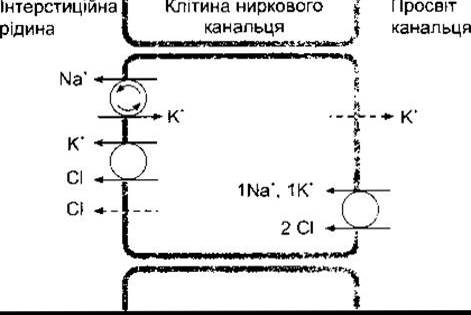
Рис. 38-16. Котранспортування Na+, К+ і Cl- шляхом вторинного активного транспортування в клітинах товстої частини висхідного коліна петлі Генле. Суцільними лініями позначено активне транспортування або вторинне активне транспортування, штриховими - дифузію. Активне транспортування за допомогою Nа+-К+-АТФ-ази позначено стрілками у колі.
Збірні трубочки
Збірні трубочки складаються з двох частин: кіркової і мозкової, через які фільтрат переходить з кори у ниркову миску. Осмотичний тиск і об’єм рідини, що міститься в збірних трубочках, залежить від кількості вазопресину, що впливає на трубочки. Цей антидіуретичний гормон, що його виділяє задня частка гіпофіза, збільшує проникність збірних трубочок до води. Цього ефекту досягають швидкою появою водних каналів з аквапорину-2 на люмінальній мембрані головних клітин (див. Розділ 14). За наявності вазопресину, достатнього для максимального антидіурезу, вода виходить з гіпотонічної рідини і надходить через кіркові збірні трубочки у міжклітинний простір кіркового шару, а рідина у збірних трубочках стає ізотонічною. За цим механізмом виходить до 10% відфільтрованої води. Далі ця ізотонічна рідина надходить у мозкові збірні трубочки; концентрація інуліну в ній у 20 разів перевищує його концентрацію в плазмі. В гіпертонічному міжклітинному просторі мозкового шару реабсорбується ще 4,7% фільтрату, внаслідок чого утворюється концентрована сеча, де концентрація інуліну перевищує його концентрацію в плазмі вже в 300 разів. Осмотичний тиск сечі у людини може досягати 1400 мосм/кг Н2O, що майже в п’ять разів перевищує осмотичний тиск плазми крові; у цьому разі реабсорбується 99,7% відфільтрованої води і лише 0,3% її з’являється в сечі (див. табл. 38-7). У інших біологічних видів здатність концентрувати сечу є навіть більшою. Максимальний осмотичний тиск сечі у собак становить близько 2500 мосм/кг, у лабораторних щурів - 3200 мосм/кг, а в деяких видів пустельних плазунів - до 5000 мосм/кг.
Без вазопресину епітелій збірних трубочок порівняно непроникний для води. Отже, рідина є гіпотонічною, і великі її кількості надходять у ниркову миску. У людини осмоляльність сечі може становити 30 мосм/кг Н2O. Непроникність дистальних відділів нефрона не є абсолютною; навіть якщо вазопресину нема, то разом з електролітами виводиться близько 2% відфільтрованої рідини. У будь-якому разі екскретується до 13% відфільтрованої рідини і діурез у цьому випадку може сягати понад 15 мл/хв (21,6 л/добу). Дані про вплив наявності і відсутності дії вазопресину на метаболізм води наведені в табл. 38-7.
Причини нецукрового діабету - захворювання, спричинюваного дефіцитом вазопресину чи недостатньою чутливістю органів-мішеней до цього гормону, - описані у Розділі 14. Наприклад, у разі нефрогенного нецукрового діабету нема реакції збірних трубочок на вазопресин. Описано дві форми цього захворювання. Перша зумовлена мутацією гена V2-рецептора, що робить його нечутливим. Ген V2- рецептора розміщений в Х-хромосомі, тип успадкування - зчеплений зі статтю, рецесивний. Друга форма спричинена мутацією аутосомного гена аквапорину-2.
Течійно-протитечійний механізм
Ефективна робота концентраційного механізму залежить від підтримання збільшуваного градієнта осмотичного тиску вздовж мозкових пірамід. Цей градієнт створюють петлі Генле, які діють як течійно-протитечійний множинний механізм, і прямих судин, що відіграють роль течійно-протитечійних обмінників.
Течійно-протитечійний механізм - це система, у якій вхідний потік на деякій відстані рухається паралельно до вихідного потоку в безпосередній близькості від нього, однак у протилежному напрямі.
Цей процес відбувається як у петлях Генле, так і в прямих судинах у мозковій речовині нирки (див. рис. 38-2).
Функція кожної петлі Генле як протитечійного помножувача залежить від активного транспортування йонів Na+ і Сl- за межі товстої частини висхідного коліна (див. вище), високої проникності тонкої частини низхідного коліна для води (див. табл. 38-8), надходження рідини від проксимального відділу канальця та її відтікання у дистальний відділ канальця. Найліпше цей процес можна зрозуміти на підставі гіпотетичної моделі послідовних кроків для досягнення рівноваги. Щоправда, in vivo ці кроки не простежуються, і рівновага підтримується в будь-який момент часу (якщо осмотичний градієнт не «вимивається»). Ці кроки відображені на рис. 38-17 для моделі кіркового нефрона без тонкої частини висхідної петлі. Спочатку уявимо ситуацію, коли осмотичний тиск уздовж низхідної та висхідної петель, а також у міжклітинному просторі становить 300 мосм/кг Н2O (див. рис. 38-17А). Тепер припустимо, що помпи у товстій частині висхідної петлі можуть випомпувати з канальцевої рідини в міжклітинну рідину 100 мосм/кг Na+ і Сl, збільшивши осмотичний тиск міжклітинної рідини до 400 мосм/кг Н2O. У відповідь на це вода з тонкої частини низхідної петлі переходить у міжклітинну рідину, зрівноважуючи осмотичний тиск міжклітинної рідини і рідини у тонкій частині висхідної петлі (див. рис. З8-17В). Проте в тонку частину висхідної петлі постійно надходить рідина з проксимального відділу канальця, осмотичний тиск якої становить 300 мосм/кг Н2O (див. рис. 38-17С), отже, зменшується градієнт, проти якого відбувається викачування Na+ і Сl-, і ще більше цих йонів надходить у міжклітинну рідину (див. рис. 38-17). Водночас до дистального відділу канальця надходить гіпотонічна рідина, а ізотонічна, та в подальшому гіпертонічна рідина надходить у товсту частину висхідного коліна. Цей процес багаторазово повторюється, що призводить у кінцевому підсумку до створення градієнта осмотичного тиску між верхньою та нижньою частинами петлі.
У юкстамедулярних нефронах, які мають значну довжину петель і тонких висхідних колін, осмотичний градієнт простежується на значній довжині, і осмотичний тиск на верхівці петлі є найбільшим. Причина цього та, що тонка частина висхідного коліна порівняно непроникна для води, але проникна для йонів Na+ та Сl. Отже, Na+ і Сl переміщаються за концентраційними градієнтами в міжклітинний простір, що зумовлює додаткове пасивне протитечійне помноження. Чим більша довжина петлі Генле, тим вищий осмотичний тиск виникає на верхівці піраміди.
Осмотичний градієнт у мозкових пірамідах не був би довго, якби Na+ і сечовина, які містяться в міжклітинних проміжках, потрапляли у кровообіг. Ці речовини є в пірамідах тому, що прямі судини відіграють роль протитечійних обмінників (рис. 38-18). Вони дифундують з кровоносних судин, які проходять до кіркового шару, у судини, що опускаються в піраміди. На противагу цьому вода дифундує з низхідних судин у фенестровані висхідні судини. Отже, електроліти та інші розчинені речовини мають тенденцію повертатись до кровоносного русла кіркового шару, а вода оминає його, внаслідок чого підтримується підвищений осмотичний тиск. Вода зі збірних трубок, також виходить по прямих судинах і вливається в загальний кровообіг. Протитечійний обмін - це пасивний процес, який залежить від руху води, і він не зміг би забезпечити Градієнт осмотичного тиску вздовж пірамід, якби процес протитечійного помноження в петлях Генле припинився.
Варто зазначити, що в петлі Генле і збірних трубочках (під впливом вазопресину) створюється дуже значний осмотичний градієнт. Власне протитечійна система уможливлює це, розподіляючи градієнт уздовж сукупності канальців загальною довжиною понад 1 см, ніж створюючи його по різні боки шару клітин лише декілька мікрометрів завтовшки. У тварин є інші приклади протитечійних систем. Один з них - теплообмін між артеріальними та венозними судинами кінцівок. Цей процес незначно простежується і в людини, однак найбільше виражений у ссавців, які живуть у холодній воді. Тепло передається з артеріальної крові, що йде до кінцівок, у венозну, яка надходить до тулуба, внаслідок чого дистальні частини кінцівок є холодними, а організм у цілому економить тепло.
Роль сечовини
Сечовина також бере участь у створенні осмотичного градієнта в мозкових пірамідах і концентруванні сечі в збірних трубочках. Її транспортують специфічні транспортери; найвірогідніше, це відбувається шляхом полегшеної дифузії. Є принаймні чотири ізоформи транспортного білка в нирках: UT-A (від UT-A1 до UT-A4), і UT-B, що містяться в еритроцитах. Сечовина рухається за межі проксимального канальця, однак, за винятком внутрішньої частини збірної трубочки, решта канальцевого епітелію непроникна для цієї речовини. Отже, внаслідок видалення води з петлі Генле та дистального відділу канальця концентрація сечовини у канальцевій рідині збільшується. Проте на рівні внутрішнього мозкового отвору збірної трубочки сечовина переходить у міжклітинну рідину ниркових пірамід, що збільшує осмотичний тиск цієї рідини. Сечовину в тій частині збірної трубочки транспортує транспортний білок UT-A1, функція якого регульована вазопресином. Із клітин нирок кролів виділено інший вазопресинозалежний транспортний білок сечовини з 10 трансмембранними доменами. Коли без впливу вазопресину осмотичний тиск канальцевої рідини на рівні внутрішньої частини збірної трубочки у мозковій речовині знижується, то сечовина переходить з міжклітинної рідини у просвіт канальця, і градієнт осмотичного тиску зменшується. Кількість сечовини в інтерстиції мозкової речовини і, відповідно, в сечі, залежить від її надходження, що, відповідно, пов’язане із кількістю спожитого білка. Отже, дієта із високим вмістом білка підвищує здатність нирок концентрувати сечу.
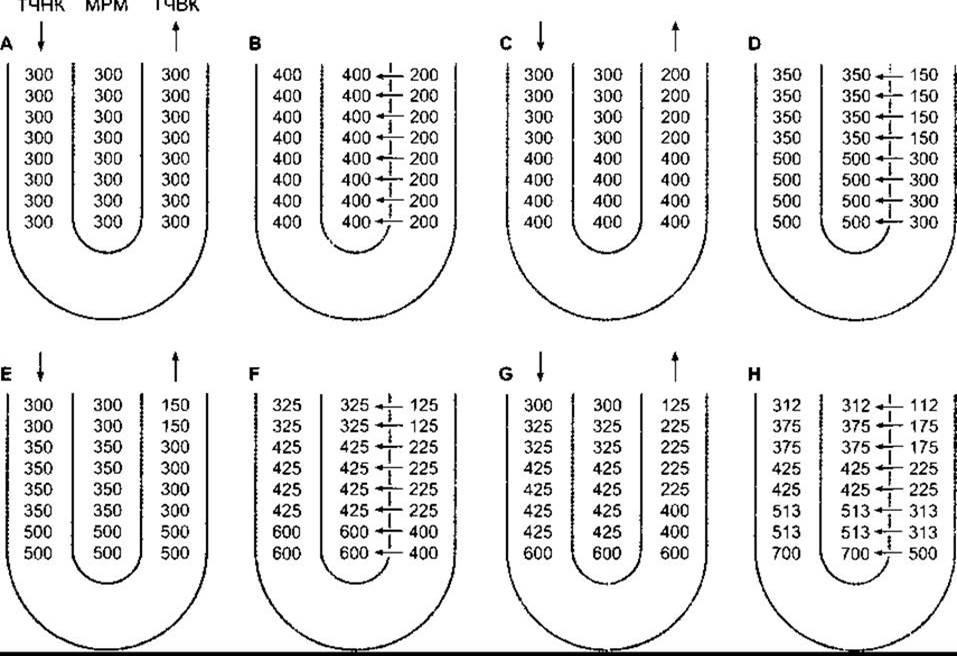
Рис. 38-17. Механізм дії петлі Генле як потокового помножувача, який створює ґрадієнт осмотичного тиску в міжклітинній рідині мозкового шару нирки (МРМ); ТЧНК - тонка частина низхідного коліна петлі Генле; ТЧВК - товста частина висхідного коліна петлі Генле. Процес створення ґрадієнта зображено у вигляді гіпотетичної послідовності кроків, починаючи з А, де осмотичний тиск в обох колінах і міжклітинній рідині становить 300 мосм/кг. Помпи, що містяться в товстій частині висхідного коліна петлі Генле, переміщають Na+ та Cl- у міжклітинну рідину, збільшуючи осмотичний тиск до 400 мосм/кг. Таким способом досягається рівновага з рідиною в тонкій частині низхідного коліна петлі Генле. Водночас продовжує надходити ізотонічна рідина в тонку частину низхідного коліна та виводиться гіпотонічна рідина за межі товстої частини висхідного коліна. Внаслідок циклічного повторення цього процесу рідина, що виходить з товстої частини висхідного коліна, стає щораз гіпотонічнішою, тоді як на верхівці петлі Генле накопичується рідина з надзвичайно високим осмотичним тиском (модифіковано та відтворено за дозволом з Johnson LR, Essential Medical Physiology, Raven Press, 1992).
Водний діурез
Механізм зворотного зв’язку, що регулює секрецію вазопресину залежно від осмотичного тиску плазми, розглянутий у Розділі 14. Після вживання значної кількості гіпотонічної рідини виділення води починається через 15 хв і досягає максимуму упродовж 40 хв. Акт ковтання рідини сам по собі призводить до незначного зменшення секреції вазопресину, однак головною причиною її зменшення є зниження осмотичного тиску плазми крові внаслідок всмоктування води.
Водне отруєння
Максимальна швидкість сечовиділення - 16 мл/хв. Якщо всмоктування рідин відбувається зі швидкістю, що перевищує це значення, то вода з гіпотонічної ПКР починає надходити у клітини, що призводить до їхнього набряку і в рідкісних випадках - до клінічно вираженого водного отруєння. Набряк клітин мозку спричинює судоми і кому, що в кінцевому підсумку призводить до смерті. Водне отруєння може виникати, якщо після введення препаратів вазопресину не було відповідно зменшено приймання рідини, або внаслідок посилення секреції вазопресину у відповідь на неосмотичні чинники, такі, наприклад, як хірургічна травма.
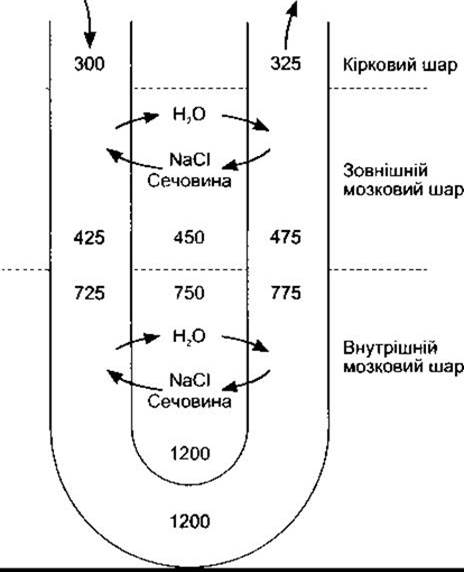
Рис. 38-18. Прямі судини і потоковий помножувач у нирці; NaCl і сечовина шляхом дифузії переходять з висхідного коліна у низхідне, тоді як вода - з низхідного коліна у висхідне.
Осмотичний діурез
Наявність у ниркових канальцях великої кількості речовин, які не піддались абсорбції, призводить до збільшення об’єму сечі, яке називають осмотичним діурезом. Речовини, які не реабсорбуються в проксимальних канальцях, чинять значний осмотичний вплив, оскільки зі зменшенням об’єму рідини їхня концентрація збільшується. Отже, вони «затримують воду» в канальцях. Треба також зазначити, що йони Na+ не можуть необмежено виводитись за межі канальця, є певний ліміт градієнта їхньої концентрації. В нормі реабсорбція води з проксимального канальця перешкоджає будь-яким відчутним змінам концентраційних градієнтів, та коли швидкість реабсорбції води зменшується, то зменшується також і концентрація Na+. Це відбувається внаслідок наявності в канальцевій рідині значної кількості різноманітних нереабсорбованих сполук. У разі досягнення граничного значення концентраційного градієнта подальша реабсорбція Na+ припиняється, і в канальці залишається більша кількість Na+ та, відповідно, затримується рідина. Внаслідок описаних вище процесів збільшується об’єм ізотонічної рідини в петлі Генле. Ця рідина має меншу концентрацію Na+, ніж вихідна, однак загальна кількість йонів Na+, що надходять у петлю Генле за одиницю часу, збільшується. В петлі Генле реабсорбція води і Na+ послаблюється, бо знижується осмотичний тиск рідини в мозковому шарі нирки. Це зниження зумовлене передусім, послабленням реабсорбції Na+, К+, і Сl у висхідному коліні петлі Генле, бо граничний концентраційний градієнт Na+ уже досягнутий. Через дистальний від діл канальця проходить більший об’єм рідини. Внаслідок зменшення градієнта осмотичного тиску вздовж ниркових пірамід у збірних трубочках реабсорбується менше води. Завдяки цьому значно збільшується об’єм сечі і виведення з нею Na+ та інших електролітів.
Осмотичний діурез зумовлюють такі речовини, як манітол (препарат групи осмотичних діуретиків) та інші полісахариди, які піддаються фільтрації, але не реабсорбуються. Це явище можуть спричинювати і деякі природні сполуки, якщо вони надходять у кількостях, що перевищують реабсорбційну здатність канальців. Наприклад, у разі цукрового діабету глюкоза, що затримується в канальцях, зумовлює поліурію, коли її фільтраційне навантаження перевищує ТмГ. Осмотичний діурез можна спричинити довенним уведенням великої кількості NaCl або сечовини.
Важливо розрізняти осмотичний і водний діурез. У випадку водного діурезу об’єм рідини, що реабсорбується в проксимальній частині нефрона, нормальний; максимальна швидкість сечовиділення становить близько 16 мл/хв. У разі осмотичного діурезу посилене сечовиділення відбувається внаслідок зменшення реабсорбції води в проксимальній частині канальця і в петлі Генле; у цьому разі може простежуватись виділення дуже великої кількості сечі. Незважаючи на максимальну здатність секреції вазопресину, концентрація сечі наближається до концентрації плазми крові (рис. 38-19), оскільки щораз більшу частину сечі становить ізотонічна рідина проксимального канальця. Якщо осмотичний діурез створюється у піддослідної тварини, яка має нецукровий діабет, то концентрація сечі, навпаки, збільшується з тієї ж самої причини.
Зв’язок концентрації сечі та швидкості клубочкового фільтрування
Значення осмотичного градієнта вздовж мозкових пірамід збільшується, якщо швидкість плину рідини через петлі Генле зменшується. Зменшення ШКФ, наприклад, унаслідок збезводнення, призводить до зменшення об’єму рідини, що надходить до течійно-протитечійного механізму, внаслідок чого зменшується швидкість плину рідини через петлі Генле і сеча стає концентрованішою. Якщо ШКФ значно зменшується, то сеча стає досить концентрованою навіть без впливу вазопресину. Якщо ж у піддослідної тварини з нецукровим діабетом частково перетиснути одну ниркову артерію, то сеча, що виділяється з цієї нирки, буде гіпертонічною внаслідок зменшення ШКФ, тоді як сеча з протилежного боку - гіпотонічною.
Кліренс вільної рідини
З метою оцінити втрати чи, навпаки, зберегти рідину внаслідок екскреції концентрованої чи розведеної сечі часто обчислюють кліренс вільної рідини (КН2О). Це різниця між об’ємом сечі та кліренсом розчинених у ній речовин (Косм):
![]()
де V - швидкість сечовиділення; Сосм і Посм - відповідно, осмотичні тиски сечі і плазми. Отже, Косм - це кількість води, що потрібна для екскреції осмотичного навантаження, яке міститься в сечі, ізотонічної до плазми крові. Значення, KH2О від’ємне, якщо сеча гіпертонічна, і додатне, якщо вона гіпотонічна. У табл. 38-8, наприклад, наведені такі дані: за максимального антидіурезу значення КН2О становить -1,3 мл/хв (-1,9 л/добу), а без дії вазопресину 14,5 мл/хв (20,9 л/добу).
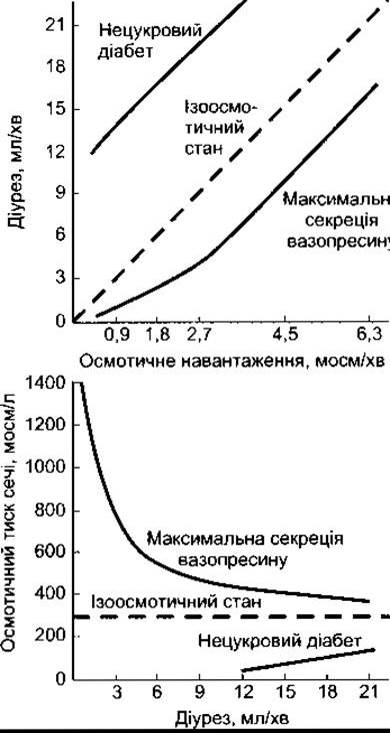
Рис. 38-19. Приблизний зв’язок між концентрацією сечі та її плином у разі осмотичного діурезу в людини. Штрихова лінія на нижньому графіку показує концентрацію, за якої осмотичний тиск сечі та плазми крові вирівнюється (відтворено за дозволом з Berliner RW, Giebisch G: Best and Taylor’s Physiological Basis of Medical Practice, 9th ed. Williams & Wilkins, 1979).