Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Зір
Відповіді зорових шляхів і кори
Шляхи до кори мозку
Аксони гангліонарних клітин забезпечують детальну просторову проекцію сітківки на бічне колінчасте тіло. Кожне колінчасте тіло має шість відокремлених шарів клітин (рис. 8-21). Шари 3-6 утворені дрібними клітинами, тому їх називають дрібноклітинними. Шари 1-2 містять великі клітини і, відповідно, їх називають великоклітинними. Шари 1,4 та 6 кожного колінчастого тіла отримують імпульси від ока протилежного боку, тоді як шари 2, 3 та 5 - від ока того ж боку. У кожному шарі є надзвичайно деталізоване відображення сітківки, і усі шість шарів перебувають у певній взаємозалежності, тобто рецепторні поля клітин, що локалізовані вздовж проведеного через шари перпендикуляра, практично збігаються. Зазначимо, що лише 10-20% імпульсів, які виходять із бічного колінчастого тіла, досягають сітківки. Значна частина додаткових імпульсів походить із зорової кори та інших ділянок мозку. Механізм зворотного зв’язку із зоровою корою має важливе значення у просторовій орієнтації та координації рухів. У сітківці розрізняють два різновиди гангліонарних клітин: великі гангліонарні клітини (або М-клітини від лат. magnus - великий), які сприймають імпульси від різних типів колбочок і пов’язані з координацією рухів та об’ємним зором, і малі гангліонарні клітини (або P-клітини від лат. parvus - малий), які розрізняють імпульси, що надходять від різних типів колбочок і забезпечують сприйняття кольору, структури та форми об’єктів. Великі клітини проектують імпульси на великоклітинні шари бічного колінчастого тіла, тоді як проекція P-клітин відбувається на дрібноклітинні шари (рис. 8-22).
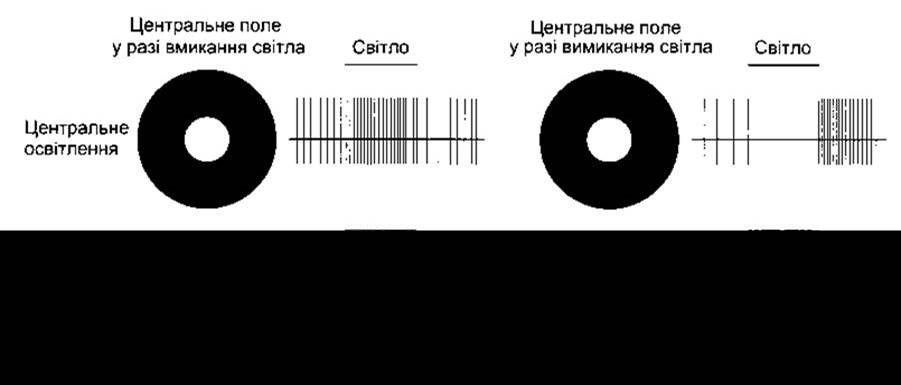
Рис. 8-20. Відповідь гангліонарних клітин сітківки на світло, що падає на частини їхніх рецепторних полів, показаних білим кольором. Біля діаграми кожного рецепторного поля наведена діаграма відповіді гангліонарної клітини, визначена шляхом позаклітинної реєстрації потенціалів дії. Зверніть увагу на те, що у трьох випадках із чотирьох інтенсивність розряду посилюється після вимкнення світла (модифіковано з Kandel Е, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000).
Від бічного колінчастого тіла велико- і дрібноклітинні шляхи проходять до зорової кори. По великоклітинних шляхах від шарів 1 та 2 (див. рис. 8-21) надходять сигнали, що стосуються сприйняття рухів, глибини і мерехтіння зображень. Дрібноклітинні шляхи передають від шарів 3-6 сигнали, що стосуються кольорів, форми і структури, а також тонких деталей об’єктів спостереження.
Клітини міжпластинчастих ділянок бічного колінчастого тіла також отримують імпульси від P-клітин, імовірно, через дендрити, що пронизують дрібноклітинні шари. Вони проектуються через окремий компонент дрібноклітинних шляхів до плямок зорової кори (див. нижче).
Первинна зорова кора
Подібно до того, як аксони гангліонарних клітин забезпечують просторове відтворення сітківки у бічному колінчастому тілі, це колінчасте тіло точка за точкою забезпечує аналогічне відтворення у первинній зоровій корі (див. рис. 8-5). З кожним нервовим волокном у зоровій корі контактує значна кількість нервових клітин. Як і інші ділянки нової кори, зорова кора складається з шести шарів (див. рис. 11- 2). Аксони клітин бічного колінчастого тіла, що формують великоклітинний шлях, закінчуються у шарі 4, а точніше, у його найглибшій частині - 4С. Багато аксонів дрібноклітинного шляху також закінчуються у шарі 4С, а аксони міжпластинчастих клітин - у шарах 2 і 3.
Шари 2 та 3 кори мозку містять скупчення клітин діаметром близько 0,2 мм, які мають вибірково високий рівень мітохондріального ензиму цитохромоксидази. Ці клітинні кластери названо плямками (блобсами, від англ. blobs) (див. рис. 8-22). Вони утворюють своєрідну мозаїку у зоровій корі і пов’язані зі сприйняттям кольорів. Дрібно
клітинний шлях також несе інформацію про зустрічні кольори до глибокої частини шару 4 (див. нижче).
Подібно до гангліонарних клітин нейрони бічного колінчастого тіла та шару 4 зорової кори відповідають на подразнення їхніх рецепторних полів збудженням центра і гальмуванням периферії або, навпаки, гальмуванням центра і збудженням периферії. Смуга світла, що покриває центр, слугує ефективним подразником, оскільки збуджує переважно центр і значно менше - периферію. Ця смуга не має орієнтаційної переваги (преференції) і як подразник однаково ефективна за будь-якого кута падіння.
Відповідь на подразнення нейронів інших шарів зорової кори суттєво відрізняється. Так звані прості клітини реагують на світлові смуги, лінії або краї лише у тому випадку, коли вони мають певну орієнтацію у просторі. Якщо, наприклад, світлова смуга повернута всього на 10° щодо преферентної орієнтації, то інтенсивність відповіді простої клітини послаблюється, а якщо кут повертання збільшити, то відповідь зникне цілковито. Є також складні клітини, подібні до простих тими перевагами, що вони надають орієнтаційну преференцію лінійним подразникам, проте водночас вони менш залежні від локалізації подразників у зоровому полі порівняно з простими клітинами або клітинами шару 4. Складні клітини звичайно генерують максимальну відповідь, якщо лінійний стимул зсунути латеральніше без зміни його орієнтації у просторі. Ймовірно, складні клітини отримують імпульси від простих клітин. Якщо у зорову кору мозку перпендикулярно до її поверхні помістити мікроелектрод і поступово заглиблювати його у різні шари, то орієнтаційна преференція нейронів буде незмінною. Отже, зорова кора, подібно до соматосенсорної (див. Розділ 7), організована у вигляді вертикальних колонок, що пов’язані з орієнтацією (орієнтаційні колонки). Кожна колонка має близько 1 мм у діаметрі. Орієнтаційні преференції сусідніх колонок упорядковано змінюються. У разі пересування від колонки до колонки орієнтаційна преференція послідовно змінюється в межах 5-10° (рис. 8-23). Тому правдоподібно, що для представництва рецепторного поля кожної гангліонарної клітини сітківки у зоровому полі кори є набір колонок у межах дрібних ділянок цього поля, що охоплюють усі можливі варіанти орієнтаційних преференцій у межах повних 360°. Прості і складні клітини називають детекторами ознак, оскільки вони сприймають і аналізують специфічні особливості того або іншого подразника. Детектори ознак виявлені також у кіркових представництвах інших органів чуття.
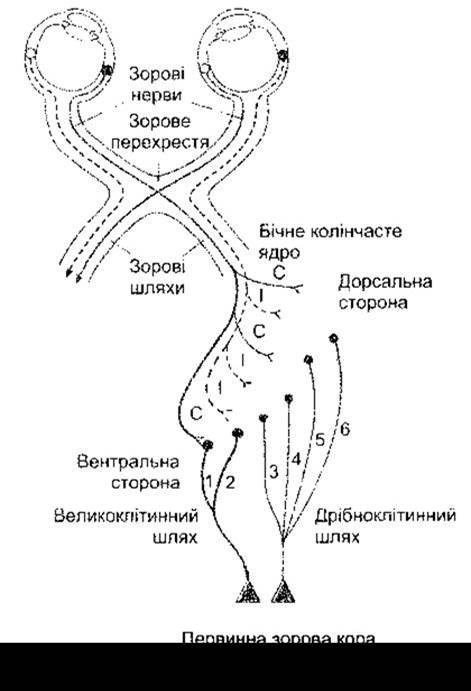
Рис. 8-21. Проекції гангліонарних клітин з правої половини сітківки кожного ока до правого бічного колінчастого тіла і далі до правої первинної зорової кори. Зверніть увагу на шість шарів колінчастого тіла. Гангліонарні P-клітини проектуються на шари 3-6, гангліонарні М-клітини - на шари 1 та 2. Іпси- (І) та контрлатеральні (С) очі проектуються на шари, що чергуються. Не показані клітини міжшарових ділянок, які проектуються через окремі компоненти дрібноклітинного шляху на плямки зорової кори (модифіковано з Kandel Е, Schwartz JH, Jessell TM [editors]: Principles of Neural Science, 4th ed. McGraw-Hill, 2000).
Орієнтаційні колонки можна картувати за допомогою радіоактивної 2-дезоксиглюкози. Засвоєння цього похідного глюкози пропорційне до активності нейронів (див. Розділ 32). Якщо це позначення застосувати для дослідження тварин, яких подразнюють рівномірно розміщеними світловими стимулами, такими як вертикальні лінії, то мозок виявляє значну кількість складно укладених рівномірно розподілених орієнтаційних колонок на великій площі зорової кори.
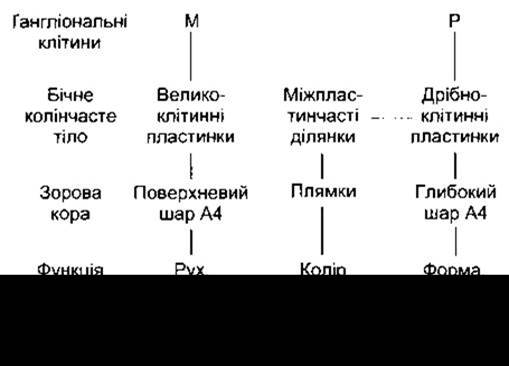
Рис. 8-22. Організація зорових шляхів. Гангліонарні М-клітини проектуються на великоклітинні пластинки бічного колінчастого ядра, а гангліонарні Р-клітини - на дрібноклітинні пластинки; P-клітини утворюють також контакти з міжпластинчастими клітинами, ймовірно, за допомогою дендритів.
Іншою особливістю зорової кори є наявність колонок домінування очей. Окремі клітини колінчастого тіла і шару 4 кори отримують вхідні імпульси лише від одного ока. Реалізація функціонування клітин шару 4 та клітин колінчастого тіла відбувається почергово. Якщо в одне око ввести велику кількість амінокислоти з радіоактивним позначенням, то ця амінокислота ввійде до складу синтезованого білка і за допомогою аксонного транспортування буде перенесена до закінчень гангліонарних клітин, а далі - через синапси у колінчастих тілах по колінчасто-шпорних волокнах до зорової кори. У шарі 4 кори позначені закінчення, що надходять від ока з радіоізотопом, чергуються із нерадіоактивними закінченнями, що надходять від іншого ока. Результатом є характерне чергування світлих і темних смужок, що охоплює значну частину зорової кори (рис. 8-24), і є незалежним від сітки орієнтаційних колонок.
Близько половини простих і складних клітин отримують імпульси від обох очей. Ці імпульси є ідентичними або близькими з огляду на охоплену зону зорової кори та преферентну орієнтацію. Однак вони відрізняються силою імпульсів, і, окрім клітин, до яких імпульси надходять лише від іпси- або контралатерального ока, є значна кількість клітин, що перебувають під впливами різного ступеня вираженості з обох очей.
Отже, первинна зорова кора розподіляє інформацію, яка стосується кольору, та інформацію про форму і рух об’єктів, інтегрує імпульси від обох очей і перетворює видимий світ у короткі відрізки різної просторової орієнтації.
Інші поля мозкової кори, пов’язані з зором
Як зазначено вище, первинна зорова кора (V1) проектується у багато різних ділянок потиличної частки та інші частини мозку. Ці ділянки часто ідентифікують номерами (V2, V3 та ін.) або літерами (LO, МТ та ін.). Локалізація деяких із цих ділянок у мозку людини показана на рис. 8- 25, а їхні ймовірні функції наведені у табл. 8-1. Функції означених ділянок досліджували у мавп, навчених виконувати різні завдання, у мозок яких пізніше були імплантовані мікроелектроди. Крім цього, використання зображень ПЕТ та функційного ЯМР (див. Розділ 16 та Додаток) уможливило виконання складних досліджень зорового пізнання та інших зорових функцій кори мозку здорових людей за умов повної свідомості. Зорові проекції з VI можна розділити на дорсальні, або тім’яні шляхи, функція яких пов’язана переважно з рухом, та вентральні, або скроневі шляхи, пов’язані з розпізнанням конфігурації, форми та облич. Окрім того, є багато перехресних зв’язків. Зорове поле V8 у людини, правдоподібно, має унікальну відповідальність за кольоровий зір.
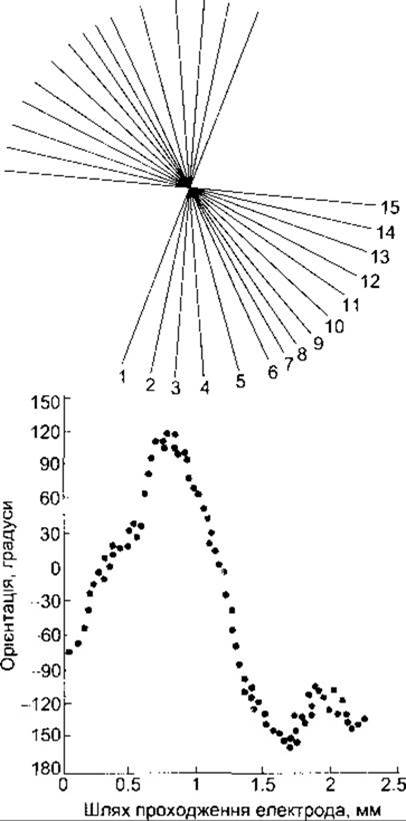
Рис. 8-23. Угорі: орієнтаційні преференції 15 нейронів, що локалізувалися на шляху косого введення електрода у зорову кору. Перевага в орієнтації змінюється поступово в напрямі проти руху годинникової стрілки. Унизу: результати аналогічного експерименту зіставлені з довжиною шляху, пройденого електродом. У цьому випадку зафіксовано численні зміни напряму ротації (модифіковано і відтворено за дозволом з Hubel DH, Wiesel TN-Sequence regularity of orientation columns in the monkey striate cortex. J Comp Neurol 1974; 158:267).
Як відомо, відбувається паралельне проходження зорової інформації по численних шляхах. Відповідно до поки що невідомого механізму вся ця інформація поступово інтегрується у сприйняття свідомого зорового образу.

Рис. 8-24. Реконструкція колонок домінування очей у сегменті шару 4 частини правої зорової кори мавпи. Темні смужки репрезентують одне око, світлі - інше (відтворено за дозволом з LeVay S, Hubel DH, WieselTN: The pattem of ocular dominance columns in macaque visual cortex revealed by reduced silver stain. J Comp Neurol 1975;159:559).