Фізіологія людини - Вільям Ф. Ґанонґ 2002
Функції нервової системи
Зір
Механізм фоторецепції
Походження електричних імпульсів
Різниця потенціалів, яка ініціює генерування потенціалу збудження, зумовлена впливом світла на світлочутливі елементи паличок і колбочок. Поглинання ними світлових променів спричинює конформаційні зміни їхніх молекул, що ініціює серію послідовних перетворень, які активують нейрони.
Око є унікальним органом, оскільки потенціали дії фоторецепторних клітин, як і електричні імпульси переважної більшості його нейрональних елементів, є локальними та ступеневими, і лише генерування потенціалів гангліонарних клітин, що здатні передаватися на значну відстань, відбувається за принципом «усе або нічого».
Відповіддю на подразнення паличок, колбочок та горизонтальних клітин є гіперполяризація (рис. 8-14), відповіддю біполярних клітин - гіпер- або деполяризація, тоді як амакринні клітини генерують потенціали деполяризації і піки, які можуть слугувати ініціаторами збудження гангліонарних клітин.
У разі порівняння потенціалів дії у колбочках та паличках простежують, що потенціал дії колбочок має гострий початок і закінчення, тоді як потенціал дії паличок має гострий початок і пологе закінчення. Криві, що узалежнюють амплітуди потенціалів дії від інтенсивності збудника, подібні у паличок і колбочок, проте палички значно чутливіші. Тому відповідь паличок пропорційна до інтенсивності збудника, якщо рівні опромінення нижчі від порогового рівня колбочок. З іншого боку, відповідь колбочок пропорційна до інтенсивності збудника у разі високих рівнів опромінення, коли відповідь паличок досягла максимуму і вже не може змінюватися. Ось чому колбочки генерують адекватну відповідь на зміни інтенсивності освітлення, вищі від певного рівня, однак погано відображають абсолютну освітленість, тоді як палички сприймають абсолютну освітленість.
Йонні основи фоторецепторних потенціалів
Натрієві канали зовнішніх сегментів паличок і колбочок відкриті у темряві, так само як постійним є потік йонів із внутрішніх до зовнішніх сегментів (рис. 8-15). Потік йонів також рухається в напрямі синаптичного закінчення фоторецепторної клітини; Nа+-К+-АТФ-аза внутрішнього сегмента підтримує йонну рівновагу. Вивільнення синаптичного нейротрансмітера стабільне у темноті. Коли світло потрапляє на зовнішній сегмент, то спричинені ним реакції закривають частину Na+-каналів, наслідком чого є гіпер поляризувальний рецепторний потенціал. Гіперполяризація зменшує вивільнення синаптичного нейротрансмітера, що, відповідно, слугує сигналом, який у кінцевому підсумку зумовлює генерування потенціалів дії гангліонарними клітинами. Ці потенціали дії надходять до мозку.
Світлочутливі речовини
Світлочутливі речовини очей людини та більшості ссавців утворені білковим компонентом, опсином, а також ретиненом, - альдегідом вітаміну А1. Термін ретинен1 використовують для вирізнення цієї речовини від ретинену2, що міститься в очах деяких видів тварин. Молекули ретиненів - це альдегіди, тому часто їх називають ретиналями. Вітаміни групи А є спиртами, і їх називають ретинолами.
Родопсин
Світлочутливий пігмент паличок називають родопсином, або зоровим пурпуром. Опсин у складі молекули родопсину називають скотопсином. Родопсин виявляє максимум чутливості до світла з довжиною хвилі 505 нм. Родопсин людини має молекулярну масу 41 000. Він локалізований у мембранах дисків паличок, становить до 90% білка цих мембран і є одним із багатьох рецепторів з покрученою формою молекули, що зв’язані з G-білками (див. рис. 1-40). Ретиналь, розміщений паралельно до поверхні мембрани (рис. 8-16) і приєднаний до залишку лізину у 296-му положенні сьомого трансмембранного домену.
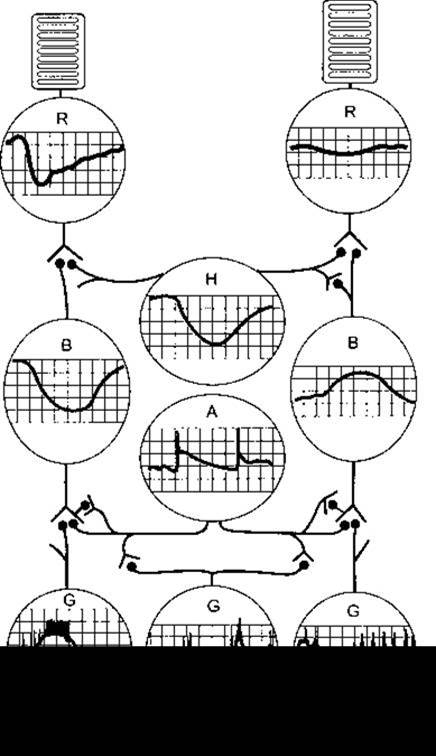
Рис. 8-14. Внутрішньоклітинна реєстрація відповіді клітин сітківки на світлове подразнення. Показані також синаптичні контакти між клітинами. Паличка (R) ліворуч сприймає спалах світла, тоді як паличка праворуч сприймає постійне освітлення помірної сили. Н - горизонтальна клітина; В - біполярна клітина; А-амакринна клітина; G - гангліонарна клітина (відтворено за дозволом з Dowling JE: Organization of vertebrate retinas. Invest Ophthalmol 1970;9:655).
У темноті ретиналь1 родопсину перебуває в 11-цис-конфігурації. Дія світла полягає у зміні форми молекули ретиналю, - її перетворенні на повний транс-ізомер (рис. 8- 17). Це, відповідно, впливає на конфігурацію опсину, зміна форми якого активує приєднаний до нього гетеротримерний G-білок, який називають трансдуцином, або Gt1. Цей білок обмінює ГДФ на ГТФ, після чого від нього відокремлюється а-субодиниця, яка зберігає притаманну їй ГТФ-азну активність доти, доки відбувається гідроліз ГТФ. Припинення дії трансдуцину прискорюється під впливом зв’язування ним ß-арестину (див. Розділ 4); а-субодиниця активує цГМФ фосфодіестеразу, яка перетворює цГМФ на 5'-ГМФ (рис. 8-18); цГМФ забезпечує утримання Na+-каналів відкритими, зменшення ж цитоплазматичної концентрації цГМФ призводить до закриття частини цих каналів. Це спричинює виникнення потенціалу гіперполяризації.
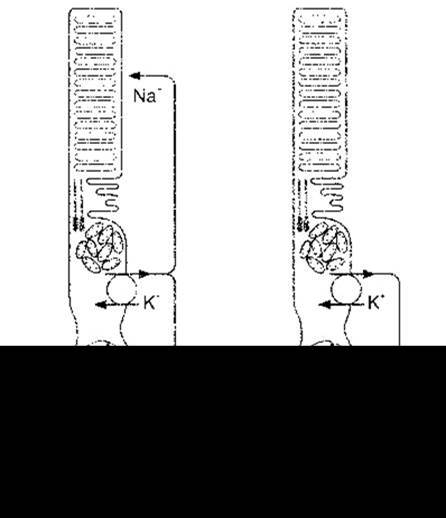
Рис. 8-15. Вплив світла на транспортування йонів у фоторецепторних клітинах. У темряві Nа+-канали зовнішнього сегмента відкриті завдяки цГМФ. Світло зумовлює перетворення цГМФ на 5’-ГМФ, що призводить до закриття частини каналів і гіперполяризації синаптичного закінчення фоторецептора.
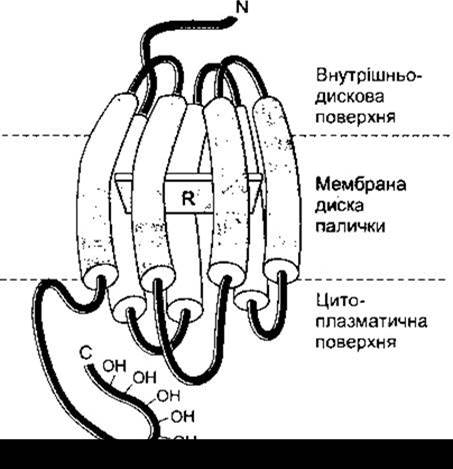
Рис. 8-16. Схематичне відтворення структури родопсину, що відображає локалізацію ретиналю1 R у мембрані диска палички.
Цикл реакцій, описаних вище, відбувається дуже швидко і посилює дію світлового сигналу. Ефект підсилення дає змогу пояснити надзвичайно високу чутливість паличкових фоторецепторів, які здатні генерувати відповідь навіть на один фотон світла.
Після набуття повної транс-конфігурації, ретиналь1 відокремлюється від опсину (явище вибілювання). Частина родопсину підлягає прямій регенерації, частина ретиналю, за наявності НАДН відновлюється ензимом алкогольдегідрогеназою до вітаміну А1, який, відповідно, взаємодіє зі скотопсином для формування родопсину (див. рис. 8-17). Усі означені реакції, за винятком утворення повного транс-ізомеру ретиналю1, не залежать від інтенсивності освітлення і відбуваються однаково добре на світлі та в темряві. Кількість родопсину у рецепторах обернено пропорційно залежить від інтенсивності освітлення.
Піґменти колбочок
У приматів є три різновиди колбочок. Ці рецептори забезпечують кольорове сприйняття зображень і максимум чутливі до світлових променів з довжиною хвилі 440, 535 та 565 нм (див. нижче). Кожен з них містить ретиналь1 та опсин. Опсин колбочок нагадує той, який належить до складу родопсину паличок. Його молекула семиразово пронизує мембрану колбочки і має свої особливості у кожного типу колбочок. Як зазначено, плазмолема колбочки, інвагінуючись, утворює мішечки, на відміну від дисків, характерних для паличок. Відповідь колбочок на світлове подразнения загалом нагадує реагування паличок. Світло активує ретиналь1, який, відповідно, активує білок Gt2, що має деякі відмінності від трансдуцину паличок; Gt2 активує фосфодіестеразу, каталізуючи перетворення цГМФ на 5'- ГМФ. Результатом є закриття Na+-каналів між позаклітинною рідиною і цитоплазмою колбочки, зменшення внутрішньоклітинної концентрації Na+ та гіперполяризація синаптичного закінчення.
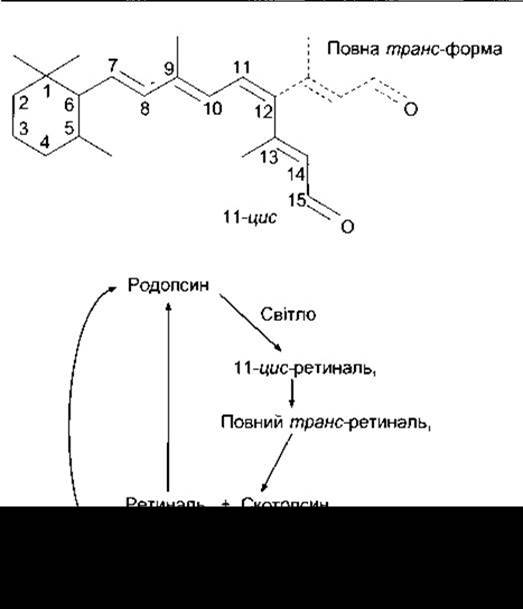
Рис. 8-17. Угорі: структура ретиналю1 з відображенням 11-цис-конфігурації (суцільні лінії) та зумовленого світлом переходу до повної транс-конфігурації молекули (штрихові лінії). Унизу: вплив світла на родопсин.
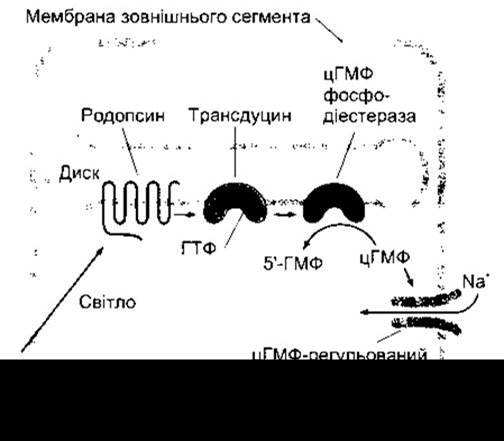
Рис. 8-18. Початкові етапи перетворення світла в паличках. Світло активує родопсин, який стимулює зв’язування ГТФ трансдуцином. Це, відповідно, активує фосфодіестеразу, яка каталізує перетворення цГМФ на 5’-ГМФ. Як наслідок, зниження внутрішньоцитоплазматичноі концентрації цГМФ призводить до закриття регульованих цГМФ йонних каналів.
Послідовність реакцій, унаслідок яких світлове подразнення фоторецепторних клітин призводить до генерування сигналу у наступному нейральному елементі сітківки, підсумована на рис. 8-19.
Ресинтез циклічного ГМФ
Світло зменшує у фоторецепторах концентрацію Са2+ і Na+. Зниження рівня Са2+ активує гуанілатциклазу, яка генерує утворення цГМФ. Вона також пригнічує активовану світлом фосфодіестеразу. Обидва означені ефекти сприяють поверненню фоторецепторів до вихідного стану шляхом відкриття Na+-каналів.
Синаптичні трансмітери сітківки
У сітківці виявлено багато різноманітних синаптичних трансмітерів, серед них ацетилхолін, глютамат, дофамін, серотонін, ГАМК, гліцин, речовина Р, соматостатин, тиротропінрилізинг гормон, гонадотропін-рилізинг гормон, енкефаліни, ß-ендорфіни, холецистокінін, вазоактивний інтестинальний поліпептид, нейротензин та глюкагон (див. Розділ 4). Каїнатні рецептори беруть участь у забезпеченні синаптичних зв’язків між колбочками та одним із різновидів біполярних клітин. Амакринні клітини єдині серед клітин сітківки синтезують ацетилхолін. У деяких видів хребетних тварин клітини, що локалізуються на межі внутрішнього ядерного і внутрішнього сітчастого шарів (див. рис. 8-2), виділяють дофамін, що дифундує по всій сітківці. Один з ефектів дофаміну виявляється у його впливі на щілинні контакти, які забезпечують вільне проходження йонних потоків між горизонтальними клітинами у темряві, збільшуючи цим розміри рецепторних полів фоторецепторів. Світло зменшує йонні потоки і роз’єднує зв’язки між горизонтальними клітинами, що досягається завдяки зростанню продукування дофаміну за умов денного світла.

Рис. 8-19. Послідовність перетворень світла в паличках і колбочках.
Формування зображень
Опрацювання зорової інформації в сітківці передбачає формування трьох зображень. Перше зображення, яке утворюється внаслідок дії світлових променів на фоторецептори, перетворюється у друге у біполярних клітинах, а те, відповідно, трансформується у третє в гангліонарних клітинах. Під час формування другого зображення сигнал модифікують горизонтальні клітини, а під час формування третього - амакринні. Зміни у разі проходження імпульсів через бічні колінчасті тіла є незначними, тому третє зображення досягає потиличних зон кори мозку.
Характерним для біполярних та гангліонарних клітин (як і для клітин бічних колінчастих тіл і клітин четвертого шару зорової кори) є те, що вони найліпше реагують на малі округлі подразники, а також те, що в межах одного рецепторного поля кільце світла навколо темного центра (окреслене освітлення) гальмує відповідь центральної точки (рис. 8-20). Центр може бути збуджувальним з гальмуванням реагування на периферії («центральноувімкнена» клітина) або гальмівним зі збудженням на периферії («центральновимкнена» клітина). Гальмування центральної відповіді прилеглими збудженими ділянками, правдоподібно, відбувається внаслідок механізму від’ємного зворотного зв’язку між сусідніми фоторецепторами, що виникає за участю горизонтальних клітин. Наприклад, активація сусідніх фоторецепторів світловим кільцем ініціює гіперполяризацію горизонтальних клітин, які, відповідно, гальмують відповідь центральних фоторецепторів. Гальмування центральної відповіді шляхом підвищення інтенсивності освітлення периферії є прикладом латерального, або аферентного, гальмування - своєрідної форми гальмування, за якої активація окремого нейрального елемента поєднується з гальмуванням прилеглих елементів. Це є загальним принципом дії сенсорних систем ссавців, який загострює сприйняття подразнень і поліпшує розпізнавання.
Електроретинограма
Електрична активність ока вивчена шляхом записування коливань різниці потенціалів між електродом усередині ока та іншим електродом на його задній поверхні або під час дослідження ока людини між поверхнею рогівки та шкірою голови. Спалах світла спричинює характерну послідовність хвиль: швидкі хвилі а та b, пов’язані з електричною активністю сітківки, та повільну хвилю с, генеровану пігментним епітелієм. Електроретинограма може допомогти в діагностиці захворювань ока у тих випадках, коли огляд сітківки неможливий з огляду на мутність середовищ ока. Цей метод обстеження також дає змогу діагностувати вроджені дистрофії сітківки, за яких офтальмоскопія виявляється неефективною.