Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Молекулярная биотехнология микробиологических систем
Генная инженерия растений: применение
Изменение окраски цветков
Цветоводы все время стараются создавать растения, цветки которых имеют более привлекательный внешний вид и лучше сохраняются после того, как их срежут. С помощью традиционных методов скрещивания за многие годы были выведены тысячи новых сортов, отличающихся друг от друга цветом и формой цветков. Однако скрещивание растений — это кропотливая процедура, требующая много времени и имеющая свои ограничения, связанные с генным пулом конкретного вида; поэтому, например, никому не удалось вывести синюю розу. В качестве альтернативы для выведения цветов с необычной окраской можно использовать методы, основанные на манипуляциях с генами ферментов биосинтеза антоцианинов. Антоцианины, соединения класса флавоноидов, являются наиболее распространенными пигментами цветков. Они синтезируются из аминокислоты фенилаланина в ходе нескольких ферментативных реакций. Окраска цветка определяется химическими свойствами их боковой цепи, при этом производные цианидина ответственны за красный цвет, а производные дельфинидина — за синий (рис. 18.15).
Дигидрофлавонол-4-редуктаза петуньи катализирует превращение бесцветного дигидрокверцетина в цианидин-3-глюкозид, соединение красного цвета, а бесцветного дигидромирицетина — в синий дельфинидин-3-глюкозид, но не может использовать в качестве субстрата бесцветный дигидрокемпферол (рис. 18.15). Однако после трансформации петуньи геном дигидрофлавонол-4-редуктазы кукурузы ее цветки приобретают кирпично-красную окраску. Этот необычный цвет, никогда ранее у петуний не наблюдавшийся, обусловлен синтезом в трансгенном растении пеларгонидин-3-глюкозида из дигидрокемпферола.
Примерно 70% объема индустрии цветоводства приходится на долю четырех растений: роз, гвоздик, тюльпанов и хризантем, поэтому все усилия по получению генетически трансформированных растений с цветками измененной окраски были направлены на работы именно с этими растениями. Например, были выведены трансгенные хризантемы, несущие смысловые и антисмысловые конструкции кДНК халконсинтазы. Этот фермент катализирует первую стадию биосинтеза антоцианина (рис. 18.15). Ученые исходили из того, что и смысловые, и антисмысловые кДНК будут подавлять экспрессию гена халконсинтазы в трансгенных растениях. «Смысловая супрессия», называемая также «косупрессией», состоит в том, что в присутствии дополнительной копии эндогенного гена подавляется накопление соответствующей мРНК. Молекулярные основы этого явления до настоящего времени не установлены. Антисмысловая же РН К халконсинтазы блокирует трансляцию эндогенной халконсинтазной мРНК. Смысловые и антисмысловые конструкции, находящиеся под контролем 358-промотора вируса мозаики цветной капусты, были встроены в бинарный вектор на основе Ті-плазмид и введены в клетки растений. У трех из 133 «смысловых» трансформантов и трех из 83 «антисмысловых» цветки были белыми, что указывало на подавление экспрессии эндогенного гена халконсинтазы, т. е. подавление синтеза антоцианина. Растения с белыми цветками вегетативно размножались черенками в полевых условиях и примерно у 90—98% из них продолжали образовываться белые, а не розовые цветки. Эта работа является важной вехой в выведении новых сортов цветов с необычной окраской, представляющих коммерческий интерес.
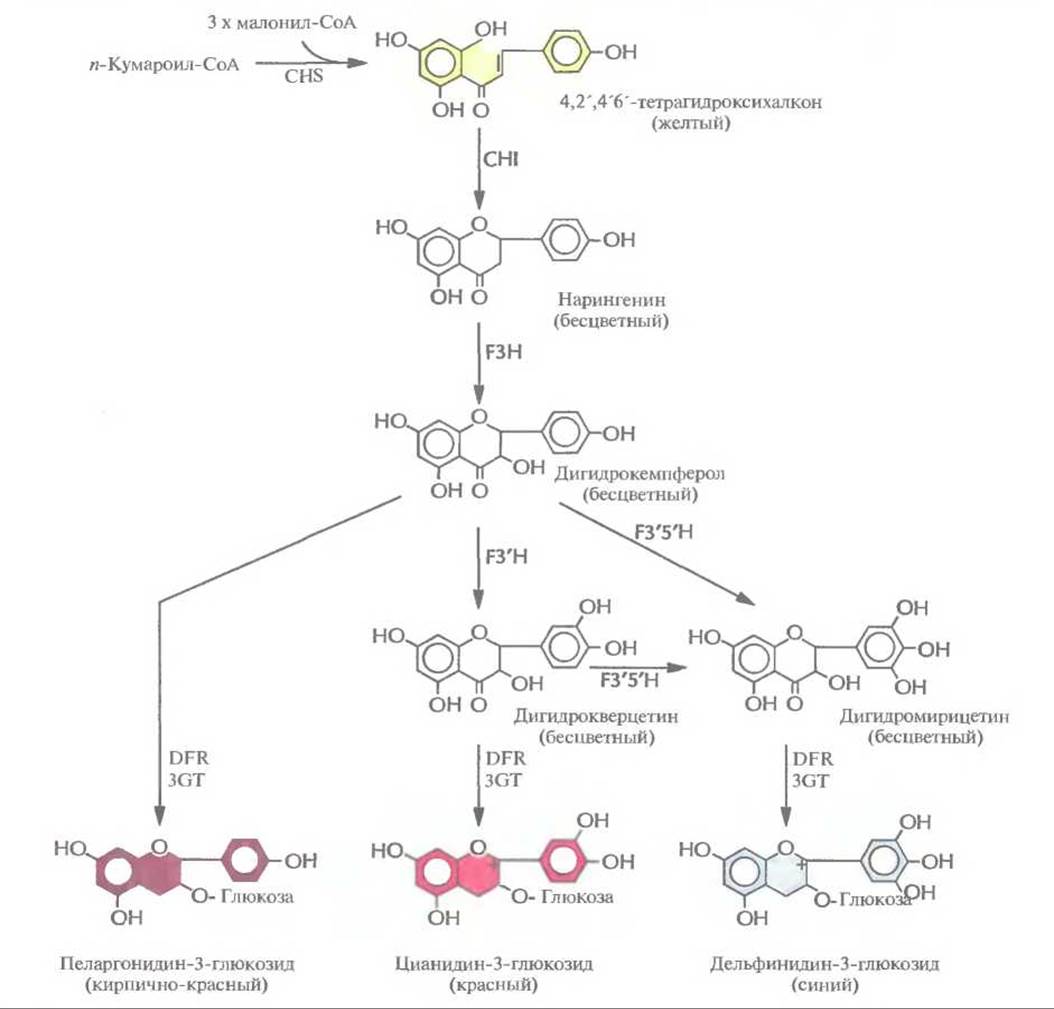
Рис. 18.15. Биосинтез антоцианинов. Сокращения: CHS — халконсинтаза, СНІ — халконизомераза, F3H — флавонон-3-гидроксилаза, F3'H — флавоноид-3'-гидроксилаза, F3'5'H — флавоноид-3'5'-гидроксилаза, DFR — дигидрофлавонол-4-редуктаза, 3GT — UDP-глюкоза: флавоноид-3-О-глюкозилтрансфераза. DFR петуньи способна катализировать превращение дигидрокверцетина в цианидин-3-глюкозид, а дигидромирицетина — в дельфинидин-3-глюкозил, соединение синего цвета. DFR кукурузы катазилирует синтез из дигидрокемпферола пеларгонидин-3-глюкозида, соединения кирпично-красного цвета.