Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Молекулярная биотехнология микробиологических систем
Генная терапия
Невирусные системы доставки генов
В опосредованной вирусами доставке генов участвуют клеточные рецепторы, с помощью которых вектор проникает в клетку-мишень, не разрушаясь лизосомными ферментами, и векторная ДНК попадает в ядро. Однако вирусные векторы имеют ряд недостатков: они дорогостоящи и часто обладают ограниченной клонирующей емкостью, что не позволяет регулировать экспрессию «терапевтического» гена с помощью тканеспецифичных последовательностей. Кроме того, вирусные белки могут вызывать воспалительную реакцию, что исключает повторное введение вектора. Поэтому были разработаны невирусные системы доставки генов.
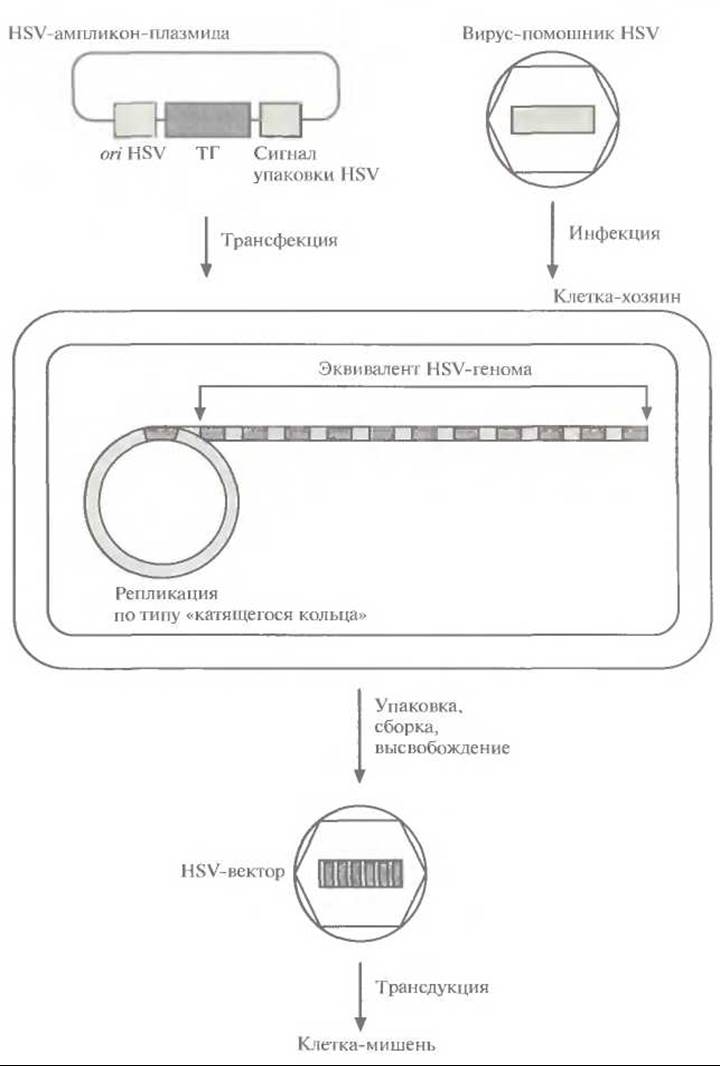
Рис. 21.9. Вектор на основе HSV-ампликон-плазмиды. Точка инициации репликации HSV (ori HSV), сигнал упаковки HSV и «терапевтический» ген (ТГ) встраивают в плазмиду Е. coli (HSV-ампликон-плазмида). Проводят трансфекцию клетки-хозяина, инфицированной вирусом-помощником HSV, полученной плазмидой. ДНК ампликон-плазмиды реплицируется по типу «катящегося кольца». 10 ампликонов, соответствующих полноразмерному геному HSV, упаковываются в HSV-капсид, который поставляет вирус-помощник HSV. Геном этого вируса не упаковывается. HSV-частицы, несущие множество копий «терапевтического» гена, высвобождаются при лизисе клетки и используются для трансдукции нейронов.
Самая простая из них — прямое введение ДНК-конструкций в клетки ткани-мишени. Если в скелетную мышцу мыши инъецировать плазмидную ДНК, то она проникнет в некоторое число клеток, о чем свидетельствует экспрессия гена-репортера в течение по крайней мере 50 сут. Однако применение этого подхода ограничивается тем, что не все ткани доступны для инъекций, а кроме того, нужны большие количества ДНК. Можно бомбардировать с помощью генного «ружья» клетки кожи или — через надрез — клетки подкожной опухоли конъюгированными с ДНК частицами золота диаметром 1—3 мкм. Введенные таким образом «терапевтические» гены экспрессируются в тканях-мишенях, а их продукты поступают в кровь. Это может облегчить доставку терапевтического белка в ткань-мишень, прямой доступ к которой затруднен. Однако большинство белков, в норме не присутствующих в крови, инактивируются или разрушаются ее компонентами. Для решения этой проблемы нужны дополнительные исследования.
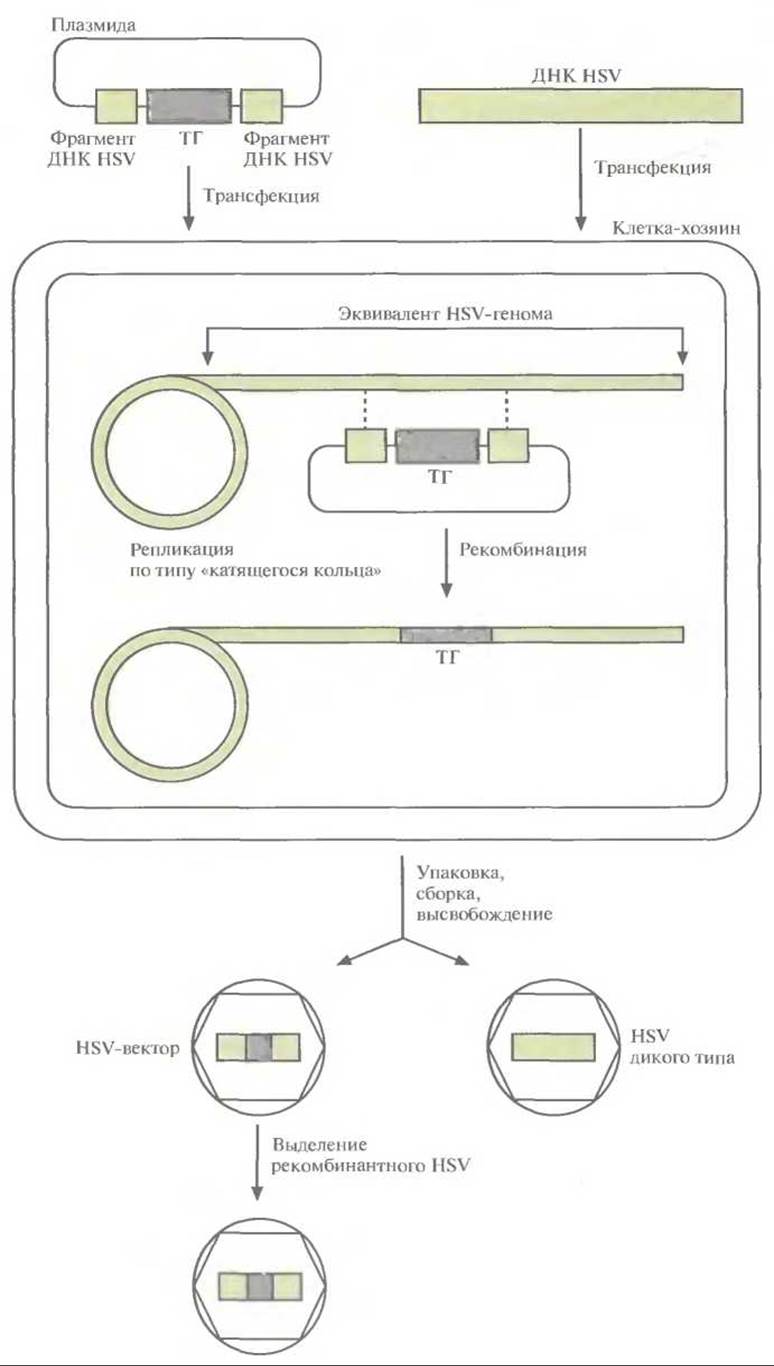
Рис. 21.10. Образование HSV-вектора с помощью рекомбинации. Проводят котрансфекцию клетки-хозяина плазмидой, которая содержит «терапевтический» ген, фланкированный последовательностями ДНК из вспомогательных областей HSV-генома, и ДНК HSV дикого типа. HSV-геном реплицируется в клеточном ядре по типу «катящегося кольца», при этом между фрагментами ДНК HSV, входящими в состав плазмиды, и ДНК HSV дикого типа может произойти рекомбинация (штриховая линия). Молекулы ДНК HSV дикого типа и рекомбинантного HSV упаковываются в вирусные частицы, высвобождающиеся из клетки после лизиса. Вирусы размножают и проводят скрининг бляшек для идентификации рекомбинантных HSV. Полученные HSV-векторы хранят в условиях, исключающих их загрязнение HSV дикого типа.
Проникновение ДНК через клеточную мембрану можно облегчить, окружив генетическую конструкцию искусственной липидной оболочкой, образующей липидную сферу (липосому) с водным содержимым. Созданы липосомы с самыми разными свойствами, например катионные липосомы, поверхность которых заряжена положительно; они связываются с отрицательно заряженной молекулой ДНК, образуя ДНК-липидный комплекс (липоплекс). Липоплексы легко образуются, относительно нетоксичны и неиммуногенны, но эффективность переноса генов с их помощью невысока, поскольку большая часть ДНК после попадания в клетку захватывается лизосомами и разрушается. Так, в одном из клинических испытаний по генной терапии муковисцидоза с использованием комплекса липосома— CFTR-тен частота трансфекции клеток назального эпителия оказалась довольно низкой, а экспрессия гена — непродолжительной.
Для доставки в клетки крупных генетических конструкций (>10 т. п. н.) с помощью эндосомного клеточного транспорта, позволяющего избежать лизосомного разрушения ДНК, образуют конъюгат ДНК с другими молекулами. Для этого полиглизин ковалентно сшивают с молекулой, связывающейся со специфическим клеточным рецептором, а затем добавляют ДНК. В результате получается компактная, плотно скрученная структура (тор), на внешней поверхности которой располагаются сайты связывания с клеточным рецептором (рис. 21.11). К сожалению, подобный конъюгат, несмотря на свою специфичность, обладает низкой эффективностью трансфекции. Все созданные к настоящему времени невирусные системы доставки имеют два основных недостатка: I) низкая частота трансфекции, не позволяющая достичь нужного терапевтического эффекта; 2) непродолжительное время экспрессии «терапевтического» гена, не обеспечивающее эффективного лечения.
Возможно, подходящим терапевтическим вектором станет искусственная хромосома человека. Это связано с: 1) возможностью включения в нее протяженных сегментов чужеродной ДНК вместе с полным набором регуляторных элементов для одного или нескольких «терапевтических» генов; 2) возможностью использования геномного варианта «терапевтического» гена, обеспечивающего высокую эффективность его экспрессии; 3) стабильностью «терапевтического» гена и его длительной экспрессией как в пролиферирующей, так и в неделящейся клетке-мишени.
Искусственная хромосома содержит три основных элемента: концевые участки (теломеры), центромеру и точки инициации репликации. Свойства теломерных областей хромосом человека хорошо изучены, чего нельзя сказать о центромерах и точках инициации репликации, и существовали опасения, что искусственную хромосому человека не удастся сконструировать, пока не будут досконально изучены все ее элементы. Однако уже получены и поддерживаются в трансфицированной культуре клеток стабильные линейные искусственные хромосомы человека (микрохромосомы), состоящие из множества ДНК-повторов (длиной около 1 м. п. н.) центромерной области Y-хромосомы, высокомолекулярных фрагментов геномной ДНК и теломерных участков. В их центромерную область был встроен ген устойчивости к неомицину, что позволило использовать среду G418 в качестве селективной. В нескольких G418-устойчивых клетках были обнаружены микрохромосомы длиной от 6 до 10 м. п. н. Две из трех микрохромосом были получены «усечением» существующей хромосомы. В одном случае исходная центромера была сохранена, а в другом заменена трансфицированной центромерной областью. Третью, полностью искусственную микрохромосому получили лигированием in vitro трех трансфицированных ДНК-элементов. Ясно, что создание искусственной хромосомы человека, содержащей «терапевтический(e)» ген(ы), вполне реально, но основной проблемой станет доставка этой огромной молекулы ДНК в ядро клетки-мишени. Кроме того, экспрессия генов, входящих в состав ДНК-блоков, из которых построена искусственная хромосома, может оказывать вредные воздействия на клетки-мишени. Для начала в ткани пациента можно попытаться имплантировать инкапсулированные клетки с искусственными хромосомами.
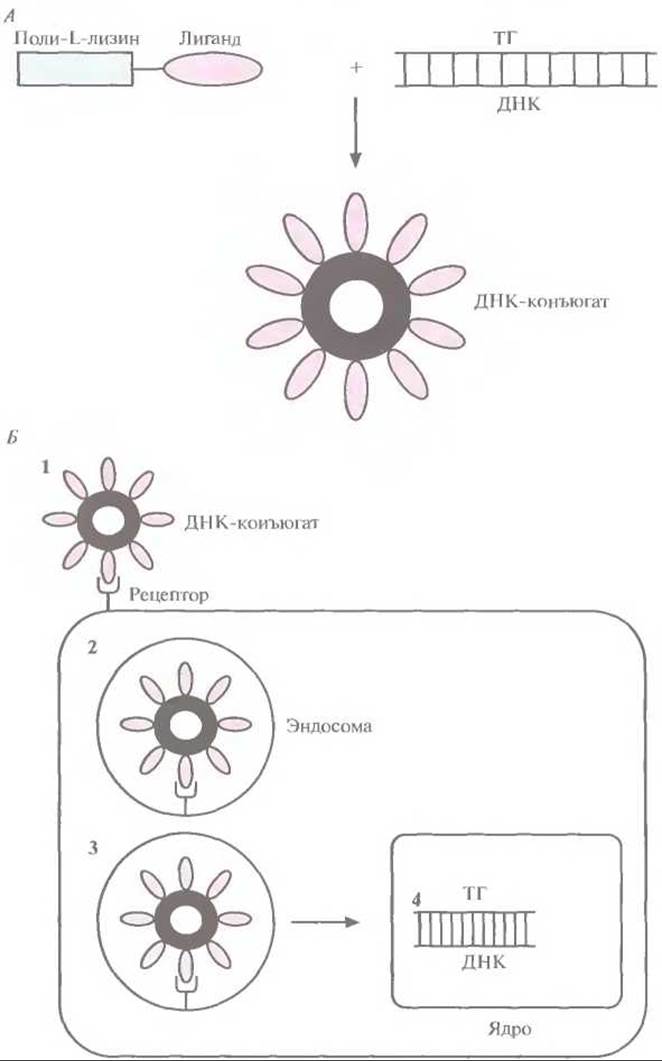
Рис. 21.11. Система доставки «терапевтических» генов с использованием ДНК-конъюгата. А. К поли-L-лизину пришивают лиганд, соединяющийся с поверхностным клеточным рецептором, и добавляют ДНК, содержащую «терапевтический» ген. В результате образуется конденсированная структура, на поверхности которой располагаются лиганды. Б. ДНК-конъюгат связывается со специфическим клеточным рецептором (1) и обволакивается клеточной мембраной (2) с образованием эндосомы (3), которая защищает его от лизосом. В эндосоме часть молекул ДНК высвобождается из конъюгата и проникает в ядро клетки (4), где и происходит экспрессия «терапевтического» гена.