Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
ДНК, РНК и синтез белка
Расшифровка генетической информации; РНК и белок
Подавляющее большинство генов содержит в закодированном виде информацию о синтезе белков. Белки — это биологические молекулы, участвующие практически во всех процессах, протекающих в живых системах. Они служат катализаторами разнообразных биохимических реакций, осуществляют транспорт веществ внутри клеток и между клетками, регулируют проницаемость клеточных мембран, из них строятся различные структурные элементы. Белки участвуют в осуществлении двигательных функций, обеспечивают защиту от инфекций и токсинов, регулируют синтез остальных генных продуктов. Основной структурной единицей белков являются аминокислоты. Все аминокислоты имеют сходное химическое строение. К центральному атому углерода (а-углерод) присоединены атом водорода (Н), аминогруппа (NH3+), карбоксильная группа (СОО-) и R-rpyппа (боковая цепь) (рис. 3.7, Л). Существует 20 разных боковых групп и соответственно 20 аминокислот. Например, в аминокислоте аланине R-группой является метальная группа (СН3). В табл. 3.1 даны одно- и трехбуквенные обозначения аминокислот. Соединяясь друг с другом пептидными связями, аминокислоты образуют полипептидную цепь. Пептидная связь образуется между карбоксильной группой одной аминокислоты и аминогруппой другой (рис. 3.7, Б). Первая аминокислота белковой молекулы имеет свободную аминогруппу (N-конец), а последняя — свободную карбоксильную группу (С-конец).
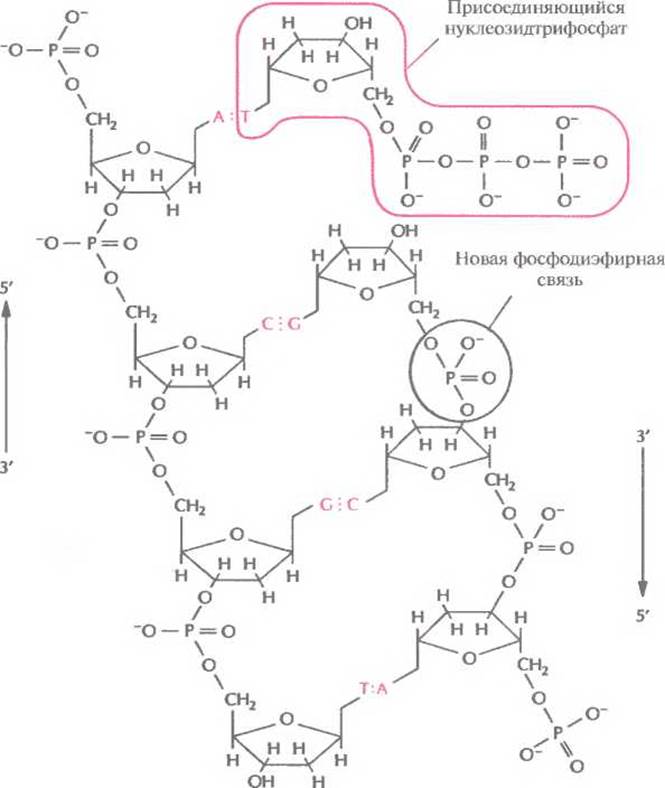
Рис. 3.6. (Продолжение) Б. Между а-фосфатной группой присоединяемого нуклеотида и 3'-гидроксильной группой растущей цепи образуется фосфодиэфирная связь. К комплементарному спариванию готов следующий дезоксирибонуклеозидтрифосфат.
Длина белковых молекул варьирует от 40 до более 1000 аминокислотных остатков, при этом в зависимости от их последовательности и от аминокислотного состава молекулы белков принимают разную форму (конфигурацию, конформацию). Многие функционально активные белки состоят из двух и более полипептидных цепей (субъединиц), как идентичных, так и несколько различающихся. Кроме того, многие белки, выполняющие ключевые функции, представляют собой сложные белковые комплексы, состоящие из множества разных субъединиц.
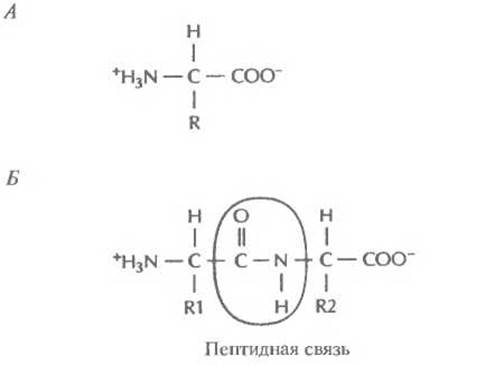
Рис. 3.7. Обобщенная структурная формула аминокислоты и образование пептидной связи. А. Аминокислота. R — боковая группа. Б. Образование пептидной связи между двумя аминокислотными остатками с боковыми группами R1 и R2.
Таблица 3.1. Аминокислоты и их обозначения
|
Аминокислота |
Трехбуквенное обозначение |
Однобуквенное обозначение |
|
Аланин |
Ala |
А |
|
Аргинин |
Arg |
R |
|
Аспарагин |
Asn |
N |
|
Аспарагиновая кислота |
Asp |
D |
|
Валин |
Val |
V |
|
Гистидин |
His |
H |
|
Глицин |
Gly |
G |
|
Глутамин |
Gln |
O |
|
Глутаминовая кислота |
Gln |
E |
|
Изолейцин |
Iso |
l |
|
Лейцин |
Leu |
L |
|
Лизин |
Lys |
К |
|
Метионин |
Met |
M |
|
Пролин |
Pro |
P |
|
Серин |
Ser |
S |
|
Тирозин |
Tyr |
Y |
|
Треонин |
Thr |
T |
|
Триптофан |
Trp |
W |
|
Фенилаланин |
Phe |
F |
|
Цистеин |
Cys |
C |
Важным «передаточным звеном» при переводе генетической информации с языка нуклеотидов на язык аминокислот являются рибонуклеиновые кислоты (РНК), которые синтезируются на определенных участках ДНК как на матрицах в соответствии с их нуклеотидной последовательностью. РНК — это линейная полинуклеотидная молекула, отличающаяся от ДНК в двух отношениях. Во-первых, моносахаридом в РНК является рибоза, содержащая не одну, а две гидроксильные группы; они связаны с 2'- и 3'-атомами углерода. Во-вторых, одним из четырех оснований в РНК является урацил (U), занимающий место тимина. Большинство молекул РНК одноцепочечные, хотя часто в них имеются взаимнокомплементарные участки, образующие двухцепочечные структуры — «шпильки» (рис. 3.8). Спаривание оснований происходит таким же образом, как и в ДНК, за исключением того, что вместо пары А—Т образуются A—U.

Рис. 3.8. Вторичная структура гипотетической молекулы РНК. Комплементарные основания соединены между собой водородными связями. Сахарофосфатный остов не изображен.
Существуют три основных типа РНК: информационная (мРНК), рибосомная (рРНК) и транспортная (тРНК). Все они играют важную роль в процессе расшифровки генетической информации. Синтез РНК на ДНК-матрице называется транскрипцией. У большинства прокариот транскрипция всех РНК осуществляется с помощью одной и той же РНК-полимеразы. У эукариот мРНК, рРНК и тРНК транскрибируются разными РНК-полимеразами.
Транскрипция во многом сходна с репликацией. Матрицей при синтезе РНК служит определенный участок одной из цепей ДНК. РНК-полимераза копирует этот участок, последовательно соединяя друг с другом с помощью 3'—5'-фосфодиэфирных связей рибонуклеотиды в соответствии с правилом комплементарности (рис. 3.9). В ходе транскрипции новосинтезированная молекула РНК отсоединяется от ДНК, и двойная спираль ДНК восстанавливается. Чтобы обеспечить транскрипцию только отдельных сегментов ДНК, должны существовать некие сигнальные последовательности, указывающие, где начинается (инициируется) транскрипция и где она останавливается (терминируется). Сигнал инициации обычно располагается перед кодирующей последовательностью, а сигнал терминации — вслед за ней. Участок ДНК, предшествующий транскрибируемому гену, называется 5'-фланкирующей последовательностью, а расположенный за ним — 3-фланкирующей. С молекулярной точки зрения ген представляет собой специфическую нуклеотидную поледовательность, транскрибируемую в РНК. Подавляющее большинство транскрибируемых последовательностей ДНК составляют так называемые структурные гены, на которых синтезируются мРНК. Конечным продуктом структурного гена является белок. У прокариот структурный ген представляет собой непрерывный участок молекулы ДНК. Транскрипция начинается со связывания РНК-полимеразы с промотором, и далее последовательно копируется весь структурный ген (кодирующая область) от первого нуклеотида до последнего с образованием функциональной мРНК (рис. 3.10). У эукариот большинство структурных генов состоит из нескольких дискретных кодирующих областей (экзонов), разделенных некодирующими областями (нитронами). По завершении транскрипции эукариотического структурного гена нитроны вырезаются из первичного продукта транскрипции с помощью ферментов, а экзоны сшиваются друг с другом «торец в торец» (сплайсинг) с образованием функциональной мРНК (рис. 3.11 и 3.12). Обычно длина экзонов составляет от 150 до 200 нуклеотидов, а длина интронов варьирует от 40 до 10 000 нуклеотидов. Очень немногие эукариотические структурные гены вообще не имеют интронов. Иногда сплайсинг мРНК может проходить по альтернативному варианту. Например, в одной ткани функциональная мРНК может образовываться в результате соединения всех экзонов первичного транскрипта, а в другой какой-то экзон будет вырезан вместе с фланкирующими его нитронами и образуется другая функциональная мРНК. Благодаря альтернативному сплайсингу в разных тканях могут образовываться разные продукты одного и того же структурного гена (рис. 3.13).
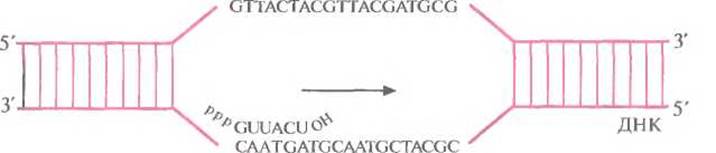
Рис. 3.9. Схематическое изображение транскрипции. Стрелкой указано ее направление.

Рис. 3.10. Схематическое изображение прокариотического структурного гена. Указаны промотор (р), сайт инициции транскрипции и ее направление (горизонтальная стрелка), область терминации транскрипции, узнаваемая РНК-полимеразой (t). Сначала на ДНК как на матрице синтезируется мРНК (транскрипция), а затем осуществляется синтез белковой цепи (трансляция).
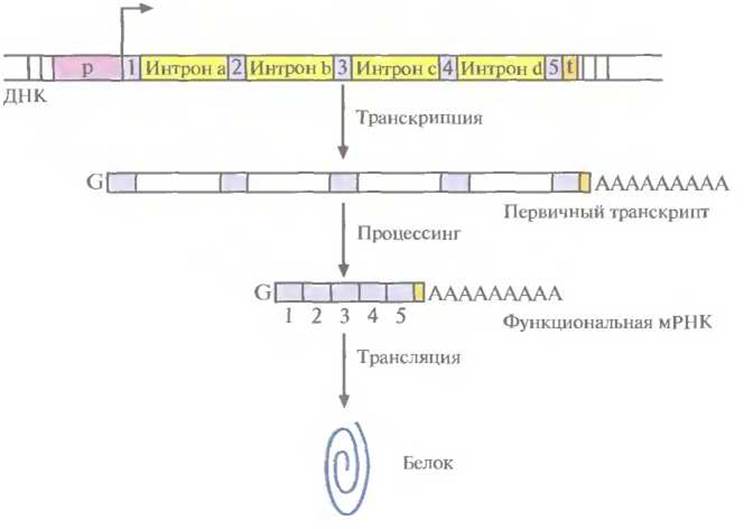
Рис. 3.11. Схематическое изображение эукариотического структурного гена. Указаны промотор (р), сайт инициации транскрипции и ее направление (горизонтальная стрелка), область терминации транскрипции, узнаваемая РНК-полимеразой (t). 1—5 — экзоны, a—d — нитроны. Первичный транскрипт содержит роlу(А)-«хвост» на 3'-конце и метилированный нуклеотид G («кэп») на S'- конце. После транскрипции интроны из первичного транскрипта вырезаются (процессинг), и на образовавшейся функциональной РНК синтезируется белковая молекула (трансляция).

Рис. 3.12. Сплайсинг первичного транскрипта у эукариот. Угловыми стрелками указаны места соединения экзонов 1, 2 и 3 после удаления нитронов а и b.
В активно функционирующей клетке примерно 3—5% суммарной РНК приходится на долю мРНК, 90% - на долю рРНК и 4% — на долю тРНК. мРНК может быть представлена десятками различных типов молекул, а рРНК — всего двумя типами. Более крупная рРНК образует с белками рибонуклеопротеидный комплекс, называемый большой рибосомной субъединицей, а рРНК меньшего размера — комплекс, называемый малой рибосомной субъединицей. Во время синтеза белков субъединицы объединяются с образованием рибосомы. У эукариот обе рибосомные субъединицы крупнее, чем у прокариот. Помимо тысяч рибосом, в клетке, активно синтезирующей белки, содержатся до 60 различных видов тРНК. тРНК — это линейная одноцепочечная молекула длиной от 75 до 93 нуклеотидов. В ней имеется несколько взаимнокомплементарных участков, спаривающихся между собой (рис. 3.8), а вся молекула укладывается в пространстве с образованием структуры, напоминающей букву L (рис. 3.14). С помощью специфических ферментов (аминоацил-тРНК—синтетаз) к 3'-концу тРНК присоединяется соответствующая аминокислота. Так, фермент аргинил-тРНК—синтетаза присоединяет к молекуле тРНКArg аминокислоту аргинин. Для каждой из двадцати аминокислот, из которых состоят все белки, существует по крайней мере одна специфическая тРНК. На другом конце молекулы тРНК расположена последовательность из трех нуклеотидов, которая называется антикодоном. Она распознает специфический кодон в мРНК и определяет, какая именно аминокислота будет присоединена к растущей полипептидной цепи.

Рис. 3.13. Альтернативный сплайсинг первичного транскрипта у эукариот. Стрелками указаны места соединения экзонов после удаления нитронов. Экзон 2, фланкируемый нитронами 1 и 2, вырезается из первичного транскрипта, а экзоны 1 и 3 соединяются с образованием функционально активной мРНК.

Рис. 3.14. Конформация тРНК, «нагруженной» аминокислотой. Штриховой прямоугольник — антикодон.