Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
Технология рекомбинантных ДНК
Плазмидные векторы
Плазмиды — это внехромосомные автономно реплицирующиеся двухцепочечные кольцевые молекулы ДНК. Плазмиды есть практически у всех бактерий. Одни из них содержат информацию, обеспечивающую их собственный перенос из одной клетки в другую (F-плазмиды), другие несут гены устойчивости к антибиотикам (R- плазмиды) или специфические наборы генов, ответственных за утилизацию необычных метаболитов (плазмиды деградации). Есть плазмиды, в которых не обнаружены гены, выполняющие какие-то определенные функции (критические плазмиды; от англ. cryptic — скрытый, латентный). Размеры плазмид варьируют от менее 1 до более 500 т.п.н. Каждая из них содержит сайт начала репликации (ori), без которого репликация плазмиды в клетке-хозяине была бы невозможна.

Рис. 4.6. ДНК-лигаза Т4 образует фосфодиэфирные связи между 5'-фосфатными и 3'-гидроксильными группами в месте разрыва в остове двухцепочечной ДНК. А. Лигирование липких концов. Б. Лигирование тупых концов.
Некоторые плазмиды представлены в клетке 10-100 копиями; они называются высококопийными. Низкокопийные плазмиды присутствуют в клетке в числе 1-4 копий. На долю плазмидной ДНК обычно приходится 0,1-5,0% суммарной клеточной ДНК. Если две или более плазмиды не могут сосуществовать в одной и той же клетке, то говорят, что они принадлежат к одной группе несовместимости. Плазмиды, относящиеся к разным группам несовместимости, беспрепятственно существуют в одной клетке, независимо от числа копий. У некоторых микроорганизмов в одной клетке было обнаружено до 8-10 разных плазмид, при этом каждая из них выполняла свои функции, была представлена характерным для нее числом копий и относилась к своей собственной группе несовместимости. Одни плазмиды несут специфичный сайт инициации репликации и могут реплицироваться только в клетках одного вида. У других плазмид этот сайт менее специфичен, и они реплицируются в самых разных бактериальных клетках. Соответственно различают плазмиды с узким и с широким спектром хозяев.
Как автономно реплицирующиеся генетические элементы плазмиды обладают всеми основными свойствами, которые позволяют использовать их в качестве вектора для переноса клонируемой ДНК. Но довольно часто природные (немодифицированные, несконструированные) плазмиды бывают лишены некоторых обязательных для «высококачественного» вектора свойств. К таким важным свойствам относятся:
1) небольшой размер, поскольку эффективность переноса экзогенной ДНК в Е. coli значительно снижается при длине плазмиды более 15 т. п. н.;
2) наличие уникального сайта рестрикции, в который может быть осуществлена вставка; 3) наличие одного или более селективных генетических маркеров для идентификации реципиентных клеток, несущих рекомбинантную ДНК. Поэтому плазмидные векторы приходится создавать с помощью генной инженерии.
Плазмидный вектор pBR322
В 80-е годы плазмидный вектор рВR322 был одним из самых популярных универсальных векторов. Обычно обозначение плазмидного вектора включает строчную букву р (от англ, plasmid) и еще несколько букв, имеющих отношение к описанию вектора или к истории его создания. Так, буквы BR в обозначении плазмиды pBR322 указывают на авторство Ф. Боливара и Р. Родригеса, сконструировавших эту плазмиду, а число 322 - цифровое обозначение, взятое из их исследовательских протоколов. Длина плазмиды pBR322 — 4361 п. н. Она несет два гена устойчивости к антибиотикам (рис. 4.7), ампициллину (Ampr) и тетрациклину (ТеР), а также уникальные сайты для BamHI, HindIII и SalI в генеТеtr, один PstI-сайт в гене Аmрr, один сайт для EcoRI, находящийся за пределами кодирующих последовательностей, и сигнал начала репликации, обеспечивающий репликацию исключительно в Е. coli. Плазмида реплицируется с образованием большого числа копий, в другие бактериальные клетки переносится с трудом. Как работает клонирующий вектор pBR322? Если очищенную кольцевую плазмиду pBR322 обработать рестриктазой, расщепляющей ее в единственном сайте, расположенном в одном из генов устойчивости к тому или другому антибиотику, то образуется линейная молекула с липкими концами. Такие молекулы смешивают с донорной ДНК, содержащей нужный ген и предварительно обработанной такой же рестриктазой. Поскольку липкие концы этих двух ДНК взаимно комплементарны, они спариваются с образованием гибридных молекул. Далее смесь обрабатывают ДНК-лигазой фага Т4 в присутствии АТР, в результате чего образуется множество разных комбинаций фрагментов, а также нежелательные продукты, в частности объединившиеся между собой фрагменты донорной ДНК и исходные плазмидные ДНК. Чтобы уменьшить количество последних, обрабатывают рестрицированную плазмидную ДНК щелочной фосфатазой, отщепляющей от линеаризованной молекулы 5'-фосфатные группы: ДНК-лигаза не может сшить концы дефосфорилированной линейной плазмидной ДНК (рис. 4,8). Что касается собственно рекомбинантных молекул ДНК, то хотя в них и имеются два одноцепочечных разрыва, ее фрагменты удерживаются вместе двумя фосфодиэфирными связями, образовавшимися с помощью ДНК-лигазы между дефосфорилированной плазмидной ДНК и рестрицированной донорной ДНК (рис. 4.8). После репликации в трансформированной клетке одноцепочечные разрывы устраняются системой лигирования клетки-хозяина.

Рис. 4.7. Генетическая карта плазмидного вектора pBR322. Гены устойчивости к тетрациклину (ТеР) и ампициллину (Amp1) содержат уникальные сайты узнавания для HindIII, SalI, BamHI и Pstl. EcoRI-сайт расположен вне этих генов. Длина вектора — 4361 п. н.
Трансформация и отбор
Теперь необходимо ввести рекомбинантную ДНК в клетку-хозяина. Этот процесс называется трансформацией. Для его осуществления используют специально разработанные приемы, например подвергают клетки высокотемпературному воздействию и обрабатывают их хлористым кальцием (СаСl2). Однако эффективность трансформации все же остается невысокой, обычно трансформируется не более одной клетки из тысячи. Таким образом, большинство клеток после проведения трансформации не содержат рекомбинантной ДНК. В некоторых из них появляется воссоединившаяся кольцевая плазмидная ДНК, избежавшая дефосфорилирования щелочной фосфатазой, в других — неплазмидная ДНК и лишь в некоторых — плазмида со встроенным фрагментом чужеродной ДНК (гибридная плазмида).

Рис. 4.8. Встраивание чужеродной ДНК в плазмидный вектор. Плазмидную ДНК, обработанную рестриктазой и щелочной фосфатазой, смешивают с рестрицированной донорной ДНК, содержащей нужный ген, и добавляют ДНК-лигазу. Два из четырех одноцепочечных разрыва при этом устраняются, и конструкция оказывается стабильной благодаря образовавшимся фосфодиэфирным связям. После введения гибридной ДНК в клетку-хозяина происходит ее репликация и образуются новые кольцевые молекулы уже без разрывов.
Как мы уже говорили, внехромосомная ДНК, не содержащая точки начала репликации, не может реплицироваться в бактериальной клетке. Таким образом, проникновение в клетку экзогенной ДНК еще не означает, что она будет поддерживаться в хозяйской клетке. Далее, для сохранения рекомбинантной ДНК в клетке-хозяине в первоначальном виде необходимо, чтобы в клетке отсутствовали гены, кодирующие синтез рестриктаз, которые могут привести к ее деградации, и чтобы клетка имела фенотип RecA - (такие клетки неспособны к общей рекомбинации, так что экзогенная ДНК не будет модифицироваться в результате гомологичной рекомбинации).
Затем необходимо идентифицировать клетки, содержащие рекомбинантную ДНК. Способ идентификации должен быть как можно более простым, поскольку приходится проверять огромное число клеток. В системе pBR322, в которой чужеродная ДНК встраивается в сайт BamHl, специфическая идентификация состоит из двух этапов. Сначала клетки после трансформации высевают на питательную среду, содержащую ампициллин. В т аких условиях могут вырасти только те клетки, в которых присутствует интактный ген Ampr — или в составе интактной плазмиды pBR322, или в составе гибридной плазмиды; нетрансформированные клетки чувствительны к ампициллину. Сайт ВаmНI расположен в гене Tetr плазмиды pBR322 (рис. 4.7); встраивание в этот ген фрагмента ДНК прерывает кодирующую последовательность, и устойчивость к тетрациклину утрачивается. Таким образом, клетки, несущие гибридную плазмиду, устойчивы к ампициллину, но чувствительны к тетрациклину, а клетки, получившие интактную плазмиду pBR322, несут ген Tetr и устойчивы как к ампициллину, так и к тетрациклину.
На втором этапе проводят разделение этих двух вариантов. Клетки, выросшие на среде с ампициллином, переносят на среду с тетрациклином методом перепечатки. Клетки, образующие колонии на чашках с тетрациклином, содержат интактную плазмиду pBR322, поскольку, как мы уже говорили, они устойчивы и к ампициллину, и к тетрациклину. Клетки, не выросшие на чашках с тетрациклином, чувствительны к этому антибиотику, значит, они содержат гибридную плазмиду pBR322.
Среди колоний, выросших на среде с ампициллином, выделяют те, которые оказались чувствительны к тетрациклину, и из каждой колонии получают индивидуальные клеточные клоны или (чаще делают именно так) объединяют все колонии, устойчивые к ампициллину и чувствительные к тетрациклину, и культивируют их вместе. Далее можно провести дополнительный скрининг и идентифицировать те клетки, которые несут гибридную плазмиду pBR322 со специфической вставкой. Присутствие сайтов HindIII и SalI в гене Tetr и сайта PstI в гене Аmрr плазмиды pBR322 позволяет изменить локализацию клонированных фрагментов чужеродной ДНК. Если для встраивания используется сайт Pstl, то отбор проводится по той же схеме, но в другом порядке, т. е. сначала высевают клетки на среду с тетрациклином, а затем — с ампициллином.
Другие плазмидные векторы
Идея использовать pBR322 как вектор для клонирования была вполне удачной, но эта плазмида содержит лишь несколько сайтов рестрикции, а отбор трансформированных клеток занимает много времени. Это привело к необходимости разработки альтернативных систем клонирования. Например, плазмида pUC19 длиной 2686 п. н. содержит: ген устойчивости к ампициллину; регулируемый сегмент гена ß-галактозидазы (lacZ') лактозного оперона Е. coli; ген lad, кодирующий репрессор, который контролирует экспрессию гена lacZ'; полилинкер — короткую последовательность с множеством уникальных сайтов узнавания для эндонуклеаз (EcoRI, SacI, Kpnl, Xmal, Smal, BamHI, XbaI, SalI, HincII, AccI, BspMI, PstI, SphI и HindIII); точку начала репликации плазмиды pBR322 (рис. 4.9).
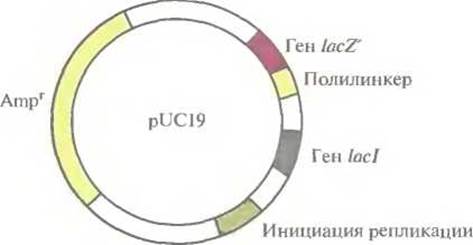
Рис. 4.9. Генетическая карта плазмидного вектора pUC19. Плазмида состоит из 2686 пар нуклеотидов и содержит уникальные сайты узнавания для EcoRI, SacI, Kpnl, XmaI, SmaI, BamHI, XbaI, SalI, HincII, AccI, PstI, BspMI, SphI и HindIII, локализованные в полилинкере; ген устойчивости к ампициллину; сайт инициации репликации, функционирующий в Е. coli; ген lad, контролирующий синтез репрессора, который блокирует транскрипцию гена lacZ' в отсутствие индуктора ИПТГ.

При отборе трансформированных клеток руководствуются следующими соображениями. Если клетки, содержащие немодифицированную плазмиду pUC19, выращивать в присутствии изопропил-β-D-тиогалактопиранозида (ИПТГ), который является индуктором lас-оперона, то продукт гена lacI не сможет связаться с промоторно-операторной областью гена lacZ', и как следствие будут происходить транскрипция и трансляция плазмидного фрагмента гена lacZ'. Продукт этого фрагмента свяжется с белком, кодируемым хромосомной ДНК, и в результате образуется активная ß-гaлактозидаза. Последовательность с множеством сайтов рестрикции (полилинкер) встроена в ген lacZ' так, что она не влияет на продукцию функциональной ß-галактозидазы, и если в среде присутствует ее субстрат 5-бром-4-хлор-3-индолил-β-О-галактопиранозид (X-Gal), то он будет гидролизоваться под действием этого фермента с образованием продукта синего цвета, окрашивающего колонии клеток, содержащих немодифицированную плазмиду pUC19.
Для клонирования в pUC19 донорную ДНК расщепляют одной из рестриктаз, чей сайт находится в полилинкере; плазмидную ДНК гидролизуют такой же рестриктазой, а затем обрабатывают щелочной фосфатазой. Обе ДНК смешивают в присутствии ДНК-лигазы Т4 и используют образовавшийся продукт для трансформации клеток, которые могут синтезировать ту часть ß-гaлактозидазы (LacZa), которая соединяется с продуктом гена lacZ' с образованием активного фермента. Обработанные клетки высевают на питательную среду с ампициллином, ИПТГ и субстратом для ß-галактозидазы. Нетрансфомированные клетки не могут расти в присутствии ампициллина, а клетки, несущие интактную плазмиду, образуют на среде с ампициллином колонии синего цвета. Клетки-хозяева, несущие гибридную плазмиду, образуют на той же самой среде белые колонии, поскольку обычно при встраивании в полилинкер чужеродной ДНК последовательности ДНК. Эту проблему можно решить, используя другую рестриктазу.
Следующий после создания библиотеки этап — это поиск клона (клонов), несущего искомую последовательность ДНК. Для этого используют три широко известных метода: гибридизацию с меченым ДНК-зондом с последующим радиоавтографическим анализом, иммунологический скрининг и скрининг по активности белка, кодируемого геном-мишенью.
Скрининг с помощью гибридизации
Нужную нуклеотидную последовательность в образце ДНК можно обнаружить с помощью ДНК-зонда, спаривающегося только с искомой последовательностью. Для этого ДНК сначала переводят в одноцепочечную форму, подвергнув ее тепловой обработке или воздействию щелочью. В этих условиях водородные связи между основаниями разрываются и цепи расходятся (происходит денатурация). Если теперь медленно снизить температуру, то произойдет их воссоединение (ренатурация). При этом, если в растворе присутствует одноцепочечный ДНК-зонд, он тоже будет ренатурировать с ДНК, специфически спариваясь с комплементарными участками. В результате образуется гибридная ДНК, т. е. двухцепочечная молекула, цепи которой принадлежат двум разным ДНК.
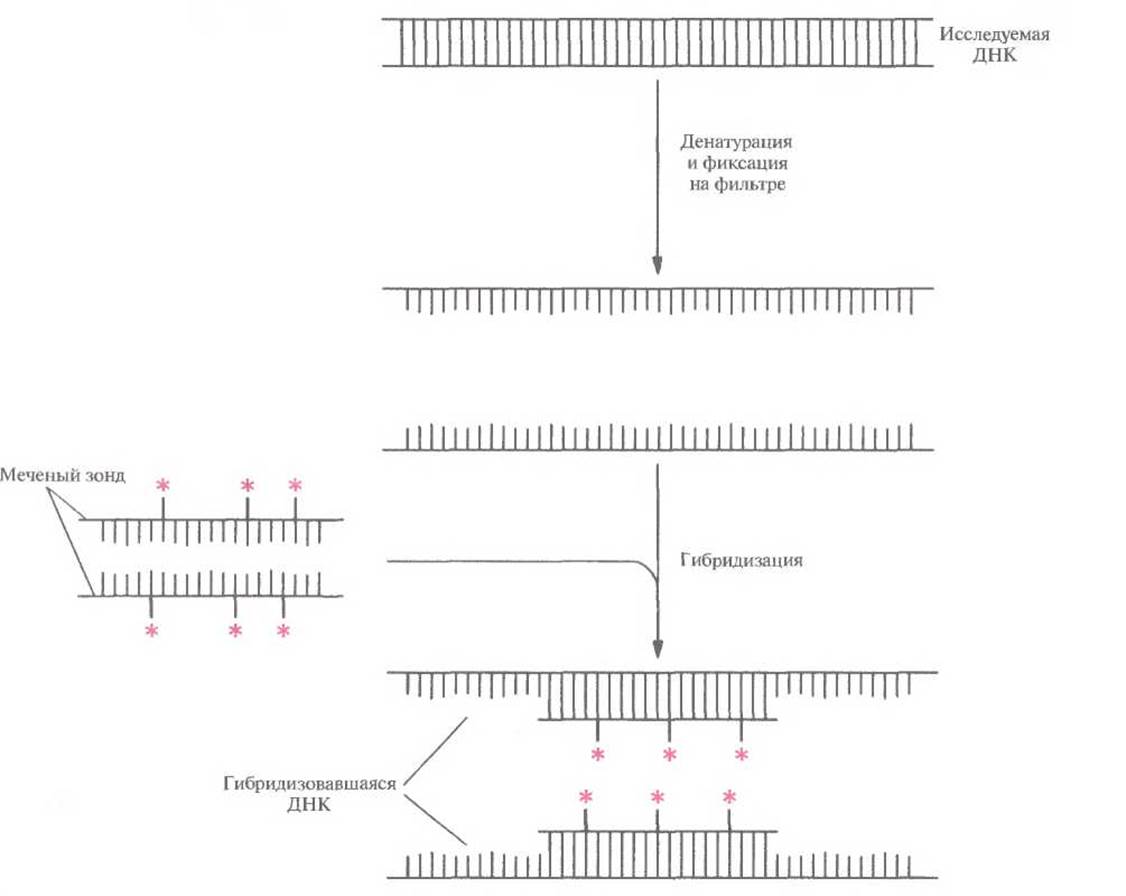
Рис. 4.11. ДНК-гибридизация. Исследуемую ДНК подвергают денатурации и фиксируют на твердой подложке, например на нитроцеллюлозном или найлоновом фильтре. Меченый ДНК-зонд (обычно длиной от 100 до 1000 п. н.) тоже денатурируют, наносят на фильтр с исследуемой ДНК и проводят их отжиг. Для удаления негибридизовавшегося ДНК-зонда фильтр промывают и визуализируют метку. Если гибридизация между зондом и исследуемой ДНК не произошла, то никакой метки на фильтре не обнаруживается. (Метка на рисунке обозначена цветной звездочкой.)

Процедура ДНК-гибридизации состоит в следующем. ДНК-мишень подвергают денатурации и одноцепочечные молекулы необратимо «пришивают» к твердой подложке (нитроцеллюлозному или найлоновому фильтру). Эту процедуру обычно проводят при высокой температуре. Затем фильтр инкубируют с одноцепочечным ДНК-зондом, меченным радиоизотопом или другой меткой. Если нуклеотидные последовательности зонда и ДНК-мишени комплементарны, то происходит их спаривание (т. е. гибридизация) (рис. 4.11). Гибридные молекулы можно визуализировать радиоавтографическим (дополнение 4.2) или другим методом, зависящим от природы метки. Если комплементарность между зондом и ДНК-мишенью отсутствует, то гибридизации не происходит, и мы получаем отрицательный результат. Обычно размер зонда варьирует от 100 до 1000 п. н. и более, хотя можно использовать как более крупные зонды, так и зонды меньшего размера. Для гибридизации, т. е. для образования стабильного комплекса, необходимо, чтобы на участке длиной 50 нуклеотидов совпадало более 80% из них, но это зависит от условий реакции.
Меченые ДНК-зонды можно получить разными способами. Один из них, называемый методом случайных праймеров, основан на применении смеси синтетических олигонуклеотидов (олигомеров), содержащих все возможные комбинации из шести нуклеотидов. Некоторые из этих олигонуклеотидов оказываются комплементарными последовательностям ДНК-мишени и гибридизуются с ними, если ДНК предварительно денатурировать (рис. 4.12). После отжига олигонуклеотидов с денатурированной ДНК-матрицей в реакционную смесь добавляют четыре дезоксирибонуклеотида (дезоксирибонуклеозидтрифосфаты; dNTP), один из них — меченый, и фрагмент ДНК-полимеразы 1 Е. coli (фрагмент Кленова). Фрагмент Кленова обладает ДНК-полимеразной и 3'-экзонуклеазной активностями, но не 5'-экзонуклеазной активностью, присущей ДНК-полимеразе I Е. coli, которая могла бы расщепить новосинтезированные молекулы ДНК. Одиночные цепи ДНК-мишени служат матрицами для синтеза новых молекул ДНК, а связанные с ними случайным образом олигонуклеотиды — затравками (рис. 4.12). При радиоактивном мечении один из dNTP содержит а-32Р, так что 32Р-меченным оказывается и сам зонд. Радиоактивную метку выявляют с помощью радиоавтографии.

Рис. 4.12. Получение меченого ДНК-зонда методом случайных праймеров. К денатурированной двухцепочечной ДНК, содержащей нуклеотидную последовательность, которую предполагается использовать в качестве зонда, добавляют гексануклеотиды (смесь всех возможных комбинаций из шести нуклеотидов) и отжигают смесь. Некоторые из олигонуклеотидов гибридизуются с немеченой денатурированной ДНК, и в присутствии фрагмента Кленова и четырех dNTP (один из которых меченый [*]) служат затравкой для синтеза комплементарной цепи. После денатурации синтезированной ДНК получают смесь меченых фрагментов ДНК, которые вместе составляют практически полноразмерную исходную ДНК-матрицу.
В качестве нерадиоизотопной метки часто используют биотин, который присоединяют к одному из четырех dNTP. Для выявления гибридизовавшегося биотинилированного зонда на фильтр наносят конъюгат стрептавидина с соответствующим ферментом (например, щелочной фосфатазой). Стрептавидин образует комплекс с биотином, который обнаруживается благодаря тому, что под действием фермента образуется окрашенное или люминеснирующее вещество — продукт превращения нанесенного на фильтр субстрата.
Зонды для скрининга геномной библиотеки можно получить по крайней мере двумя способами. Во-первых, можно использовать клонированную ДНК близкородственного организма (гетерологичный зонд). В этом случае условия гибридизации нужно подбирать таким образом, чтобы она могла происходить при существенном расхождении между нуклеотидными последовательностями зонда и искомой ДНК; это позволяет решить проблемы, связанные с заведомым различием между ДНК — источником зонда и исследуемой ДНК. Во-вторых, зонд можно получить методом химического синтеза, основываясь на известной аминокислотной последовательности белкового продукта искомого гена.
Скрининг библиотек геномных ДНК обычно проводят по следующей схеме. После трансформации высевают клетки на чашки с питательной средой и переносят выросшие колонии на твердую подложку (например, на нитроцеллюлозный или найлоновый фильтр); проводят лизис клеток, затем депротеинизацию и денатурацию ДНК; фиксируют ДНК на подложке. Наносят на фильтр меченый зонд и проводят отжиг, а затем радиоавтографию. Колонии на исходной чашке, которые содержат гибридизовавшуюся ДНК, выделяют и культивируют (рис. 4.13). Поскольку большинство библиотек получается в результате частичного гидролиза, положительный гибридизационный сигнал может быть получен для нескольких колоний (клонов). Теперь необходимо определить, какой именно клон (если таковой имеется) содержит искомый ген целиком. С помощью гель-электрофореза и картирования определяют размер каждого фрагмента (вставки) и идентифицируют аналогичные фрагменты или фрагменты с перекрывающимися последовательностями. Можно также провести дополнительное клонирование с тем, чтобы составить полный ген из перекрывающихся фрагментов. Или, если вставка в каком-либо из клонов достаточно велика и вполне может содержать весь ген, провести ее секвенирование и убедиться в наличии старт- и стоп-кодонов и полноразмерной нуклеотидной последовательности, кодирующей искомый белок.
К сожалению, никто не может дать гарантии, что в библиотеке представлена вся нуклеотидная последовательность нужного гена. Если поиск полноразмерного гена оказался безрезультатным, можно создать другую библиотеку, используя другую рестриктазу, и провести скрининг с помощью исходного зонда или зондов, созданных на основе предыдущей библиотеки. Чтобы повысить вероятность присутствия в библиотеке полной версии искомого гена, можно также создать библиотеки, содержащие фрагменты ДНК заведомо большего размера, чем средний размер прокариотического гена (этот вариант мы рассмотрим в данной главе позже).
Иммунологический скрининг
В отсутствие ДНК-зонда для скрининга геномной библиотеки можно использовать другие методы. Например, если клонированный ген экспрессируется, то его продукт — весь белок или его часть — можно обнаружить иммунологическими методами. Технически эта процедура имеет много общего с гибридизацией. Все клеточные линии (клоны) библиотеки высевают на чашки с питательной средой. Выросшие колонии переносят на фильтр, клетки лизируют, а высвободившиеся белки фиксируют на фильтре. Затем на фильтр наносят первые антитела, которые специфически связываются с данным белком (антигеном), все несвязавшиеся антитела удаляют, а фильтр помещают в раствор вторых антител, специфичных в отношении первых антител. Во многих тест-системах используют конъюгаты вторых антител с ферментом, например с щелочной фосфатазой. После отмывания фильтра добавляют бесцветный субстрат. Если вторые антитела связываются с первыми, то под действием фермента происходит гидролиз субстрата с образованием окрашенного вещества в том месте, где идет реакция (рис. 4.14).
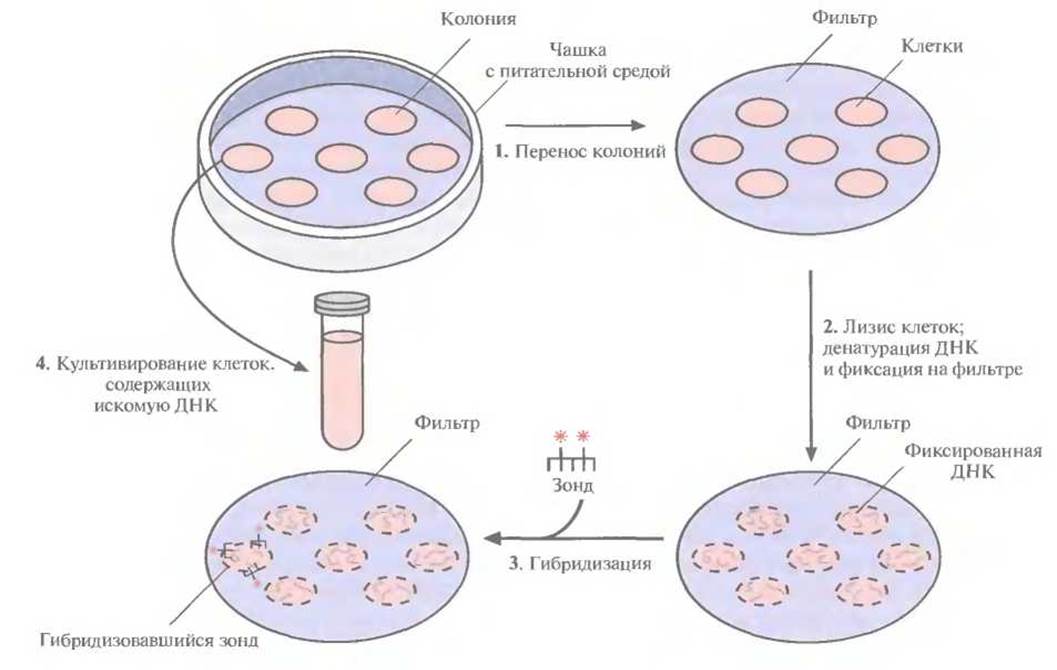
Рис. 4.13. Скрининг библиотеки геномной ДНК с применением меченого зонда. Клетки после трансформации высевают на твердую питательную среду, обеспечивающую рост только трансформированных клеток. 1. Переносят клетки из каждой выросшей колонии на твердую подложку (например, нитроцеллюлозный или найлоновый фильр) так, чтобы их расположение соответствовало таковому на чашке. 2. Клетки лизируют, высвободившуюся ДНК подвергают денатурации и депротеинизации и фиксируют на фильтре. 3. На фильтр наносят меченый ДНК-зонд и проводят гибридизацию. Смывают с фильтра негибридизовавшийся зонд и проводят радиоавтографию с тем чтобы определить, какие клетки содержат меченый ДНК-зонд. 4. Идентифицируют на чашке колонии, содержащие искомую ДНК (положительный гибридизационньгй сигнал), отбирают из них материал и культивируют.
Те клетки на чашке, которые соответствуют окрашенным пятнам на фильтре, содержат или полноразмерный ген, или достаточно протяженный его участок, обеспечивающий синтез белкового продукта, узнаваемого первыми антителами. По окончании иммунологического скрининга геномной библиотеки необходимо определить, какой именно из отобранных клонов содержит полноразмерный ген.

Рис. 4.14. Иммунологический скрининг геномной библиотеки (иммунологическое тестирование колоний). Клетки после транстформации высевают на твердую питательную среду, обеспечивающую рост только трансформированных клеток. 1. Переносят клетки из каждой выросшей колонии на твердую подложку (например, нитроцеллюлозный или найлоновый фильтр) так, чтобы их расположение в точности соответствовало таковому на чашке. 2. Клетки подвергают лизису, белки фиксируют на фильтре. 3. Наносят на фильтр первые антитела, которые связываются только с искомым белком. 4. Несвязавшиеся первые антитела удаляют, наносят на фильтр вторые антитела, специфичные в отношении первых антител, связанные с ферментом (например, щелочной фосфатазой). 5. Смывают с фильтра все несвязавшиеся вторые антитела и наносят на него бесцветный субстрат, при гидролизе которого образуется окрашенный продукт. Гидролиз может произойти только в присутствии вторых антител. 6. Отбирают колонии на чашке, соответствующие окрашенным пятнам на фильтре, и культивируют их. В них может содержаться рекомбинантная ДНК, кодирующая белок, гомологичный первым антителам.
Скрининг по активности белка
Метод гибридизации ДНК и иммунологические методы позволяют идентифицировать многие гены и их продукты. Если при этом искомый ген кодирует фермент, не синтезируемый клеткой- хозяином, то для обнаружения клонов, содержащих данный ген, можно использовать метод идентификации на чашках. Так были идентифицированы гены а-амилазы, эндоглюканазы и ß-галактозидазы различных организмов. Для этого клоны Е. coli, составляющие геномную библиотеку данных организмов, высевали на чашках с питательной средой, содержащей специфический субстрат. После окрашивания селективным красителем клетки, способные утилизировать этот субстрат, приобретали характерную окраску.
Если искомый ген кодирует продукт, без которого мутантная клетка-хозяин не может расти на минимальной среде, то библиотеку можно создать методом трансформации мутантных клеток. Клетки, выросшие в отсутствие необходимого субстрата на минимальной среде, обязательно содержат функциональный искомый ген, попавший в клетку в составе плазмидного вектора. В разных вариантах этот подход использовали для выделения многих важных генов, в частности генов, ответственных за синтез антибиотиков и образование азотфиксирующих клубеньков на корнях некоторых растений.