Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
Технология рекомбинантных ДНК
Клонирование структурных генов эукариот
Для клонирования эукариотических структурных генов необходимы специальные методики. Прокариоты не способны удалять интроны из первичных РНК-транскриптов, поэтому правильная трансляция эукариотических мРНК в бактериальной клетке невозможна. Более того, экспрессия эукариотической ДНК может осуществляться только при наличии прокариотических сигнальных последовательностей, регулирующих транскрипцию и трансляцию. Концевые участки эукариотических мРНК особым образом модифицированы: их 5'-концы кэпированы (содержат «кэп» из остатка G, часто метилированного), а 3'-концы полиаденилированы (содержат ро1у(А)-«хвост» из примерно 200 остатков аденозина).
Наличие poly (А)-хвоста позволяет отделить мРНК от рибосомной и транспортной РНК. Для этого суммарную эукариотическую РНК пропускают через колонку, заполненную целлюлозой, к которой «пришиты» короткие олигонуклеотидные цепочки из тимидиновых остатков длиной примерно 15 звеньев, oligo(dT). Poly(A)-хвосты молекул мРНК спариваются с oligo(dT) и задерживаются в колонке, а молекулы тРНК и рРНК свободно проходят через нее. Затем колонку промывают буфером, в котором происходит разрыв водородных связей между А и Т, и мРНК высвобождается.
Саму мРНК нельзя встроить в ДНК-вектор, сначала на ней необходимо синтезировать двухцепочечную ДНК. Для этого последовательно используют две разные полимеразы: обратную транскриптазу и фрагмент Кленова ДНК-полимеразы I (рис. 4.15). Вначале в реакционную смесь с очищенной мРНК добавляют короткие oligo(dT), обратную транскриптазу и четыре dNTP (dATP, dTTP, dGTP, dCTP). Роlу(А)-хвост мРНК спаривается с oligo(dT), несущим свободную 3'-ОН-группу, которая инициирует синтез комплементарной цепи. Матрицей в этом синтезе служит молекула мРНК, а катализирует его обратная транскриптаза, продуцируемая некоторыми РНК-вирусами. Она последовательно присоединяет к растущей цепи остатки Т, С, G или А, комплементарные A, G, С или U мРНК. In vitro синтез ДНК идет не до конца, при этом обратная транскриптаза перед остановкой обычно «поворачивает вспять» и присоединяет несколько нуклеотидов в обратном направлении (рис. 4.15), так что в результате образуется «шпилька».
В реакционную смесь добавляют фрагмент Кленова ДНК-полимеразы I Е. coli, который достраивает вторую цепь ДНК, используя первую цепь как матрицу. Он присоединяет дезоксирибонуклеотиды к растущей цепи, начиная с 3'-ОН-конца шпильки. По окончании синтеза препарат обрабатывают ферментом РНКазой Н, которая разрушает молекулы мРНК, и нуклеазой Sl, отщепляющей одноцепочечные концы ДНК. Полученный препарат представляет собой смесь частично и полностью двухпепочечных комплементарных ДНК-копий (кДНК) мРНК, преобладающей в исходном образце. Разные кДНК можно встроить в плазмидный вектор и получить кДНК-библиотеку. Для скрининга кДНК-библиотеки с целью идентификации клонов, несущих специфические гибридные плазмиды, можно использовать метод гибридизации или иммунологические методы. В последнем случае кДНК должна быть встроена в сайт, находящийся под контролем бактериального промотора, обеспечивающего транскрипцию. Однако практически ни один вектор не гарантирует, что во встроенной кДНК сохранится правильная рамка считывания и синтезируется правильная полипептидная цепь. Тем не менее все положительные клоны, выявленные тем или иным методом, необходимо подвергнуть дальнейшей проверке и идентифицировать те из них, которые несут полноразмерную нуклеотидную последовательность, кодирующую белок-мишень.
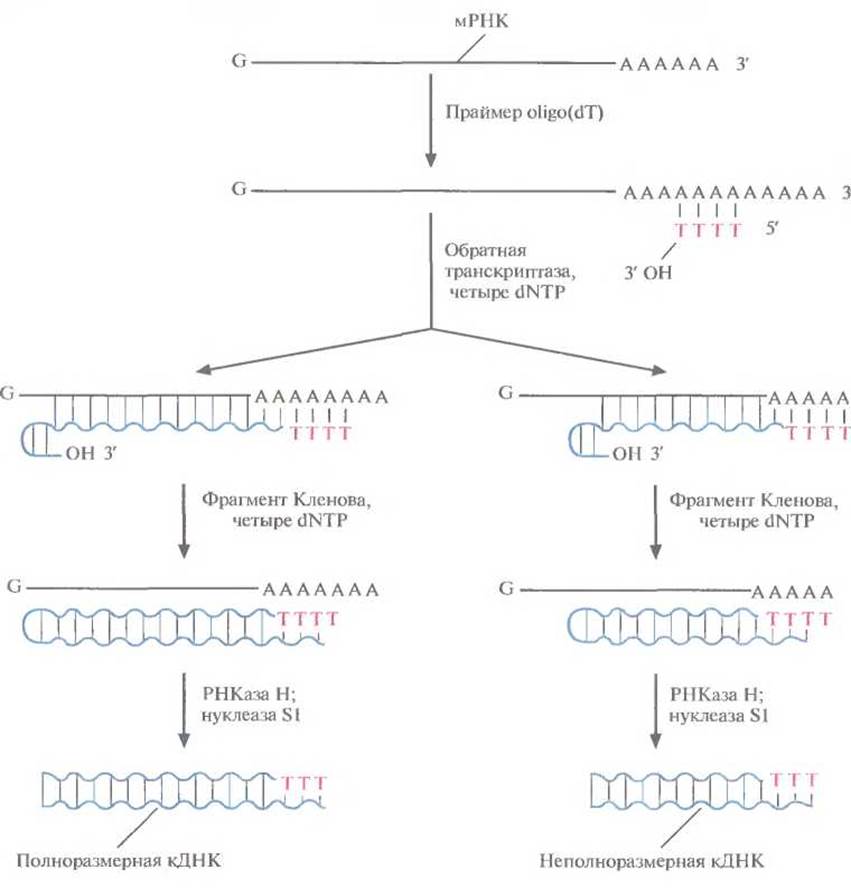
Рис. 4.15. Синтез кДНК. К препарату очищенной мРНК добавляют праймер oligo(dT). Для синтеза ДНК на РНК-матрице используют фермент обратную транскриптазу и четыре dNTP. In vitro обратная транскриптаза не обеспечивает синтез полноразмерных кДНК-копий на всех матрицах и образует на конце растущей цепи шпильку со свободной 3'-ОН-группой. Эта группа инициирует синтез второй цепи ДНК при участии фрагмента Кленова. После завершення синтеза молекулы мРНК гидролизуют РНКазой Н, а ДНК обрабатывают нуклеазой Sl, в результате чего получаются линейные молекулы ДНК с тупыми концами без шпилек.