Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Основы молекулярной биотехнологии
Получение рекомбинантных белков с помощью эукариотических систем
Другие дрожжевые системы экспрессии
С помощью систем экспрессии S. cerevisiae удалось получить много разных рекомбинантных белков. К сожалению, в большинстве случаев уровень их экспрессии был довольно низким. Кроме того, обнаружились и другие проблемы.
✵ При увеличении масштабов системы часто происходит потеря плазмид, даже если используются индуцируемые промоторы.
✵ Гетерологичный белок зачастую оказывается гипергликозилированным и содержит более 100 остатков маннозы в каждой боковой олигосахаридной цепи, в то время как в нативных белках их содержится только от 8 до 13 на цепь. Наличие лишних маннозных остатков может изменять биологическую активность продукта или его иммуногенность.
✵ Во многих экспериментах белки, которые должны были секретироваться, на самом деле концентрировались в периплазматическом пространстве, что еще более осложняло их очистку.
Все это заставило ученых исследовать возможность получения гетерологичных белков с помощью других видов дрожжей и с использованием эукариотических систем. В частности, изучались соответствующие векторы — системы экспрессии, содержащие видоспецифичные регуляторные последовательности транскрипции и трансляции, возможность трансформации этих видов и получения высокого выхода белков и возможность крупномасштабного культивирования организма-хозяина. В качестве альтернативы S. cerevisiae можно использовать Kluyveromyces lactis, дрожжи, которые применяют для промышленного производства лактозы (ß-галактозидазы); Schizosaccharomyces рот be, дрожжи, размножающиеся делением, а не почкованием; Yarrowia lipolytica, которые используют алканы в качестве субстрата; Pichia pastoris и Hansenula polymorpha, которые могут использовать метанол как единственный источник углерода и энергии.
Синтез поверхностного антигена вируса гепатита В
Метилотрофные дрожжи Р. pastoris можно без труда и больших затрат выращивать в промышленных биореакторах. Их использование в качестве организма-хозяина позволило бы увеличить выход активных продуктов — гетерологичных белков. Такой вывод можно сделать, рассмотрев в качестве примера получение поверхностного антигена вируса гепатита В (HBsAg) с помощью специально разработанной системы с использованием интегрирующего вектора. Сначала ген HBsAg встроили между промотором гена алкогольоксидазы 1 (AOXlp) и сигналом терминации-полиаденилирования (ЛОХIt) того же гена (рис. 7.5). Регуляция активности гена АОХ1 Р pastoris осуществляется с помощью метанола. В его присутствии на долю алкогольоксидазы может приходиться до 30% всех белков клетки, а в отсутствие метанола алкогольоксидаза не синтезируется вообще.
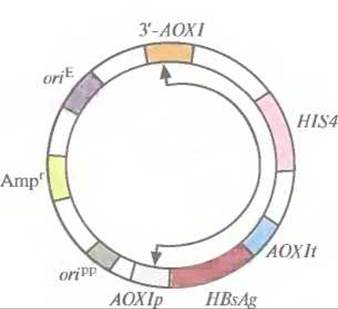
Рис. 7.5. Интегрирующий экспрессирующий вектор для Р. pastoris. Между промотором (AOXlp) и сигналом терминацииполиаденилирования (AОХlt) гена алкогольоксидазы 1 Р. pastoris встроен ген HBsAg. HlS4 — ген, кодирующий один из ферментов биосинтеза гистидина, гистидинолдегидрогеназу. Кроме того, вектор содержит сайт инициации репликации Р. pastoris (оriрр), ген устойчивости к ампициллину (Аmрr) и сайт инициации репликации, активный в Е. coli (oriE). 3'-AOXl — это фрагмент 3'-концевой последовательности гена алкогольоксидазы 1 Р. pastoris. Стрелками указан сегмент, который интегрируется в геном Р. pastoris.
Вектор (рис. 7.5), специально сконструированный для этих исследований, содержал следующие элементы: 1) блок AOXlp-HBsAg-AOXlt, 2) сайт инициации репликации, функционирующий в Р. pastoris; 3) фрагмент ДНК, содержащий сайт инициации репликации плазмиды pBR322 и селективный маркер Е. coli; 4) фрагмент 3'- АОХl, способствующий интеграции клонированной ДНК в определенный сайт хромосомы; 5) активный ген гистидинолдегидрогеназы (HIS4), кодирующий фермент, который участвует в синтезе аминокислоты гистидина. Наличие в этой конструкции последовательностей pBR322 позволяет использовать для работы с ней Е. coli, что облегчает клонирование и при необходимости позволяет получать большие количества векторной ДНК.
Чтобы предотвратить утрату плазмиды, была предусмотрена интеграция участка AOXlp-HBsAg-AOXlt в геном Р. pastoris. Для этого штамм Р. pastoris HIS4 с дефектным геном гистидинолдегидрогеназы трансформировали фрагментом вектора, содержащим элементы AOXlp-HBsAg-AOXlt, HIS4 и 3'-АОXI (рис. 7.5). В результате двойного кроссинговера между AOXlp и 3'-АОХl введенной ДНК, с одной стороны, и комплементарными последовательностями хромосомной ДНК, с другой, произошла интеграция последовательностей AOXlp-HBsAg-АОХlt и HIS4 в геном, сопровождающаяся утратой хромосомного гена АОХ1 (рис. 7.6). Клетки, в геном которых включился ген HIS4, растут на среде без гистидина; этот признак может использоваться для их отбора. Вторым критерием отбора служит замедление роста клеток в присутствии метанола, поскольку после потери гена АОХl после двойного кроссинговера активным остается только один, менее эффективный ген АОХ2.
Клон с интегрировавшим фрагментом AOXlp-HBsAg-AOXlt при росте в присутствии метанола, который активирует АОХl-промотор, синтезировал в больших количествах аутентичный белок HBsAg, накапливающийся в цитоплазме. Белковый продукт образовывал такой же мультисубъединичный комплекс, как и соответствующий белок в клетках человека, инфицированных вирусом гепатита В, и связывался с антителами к этому вирусу. При выращивании данного клона в 240-литровом ферментере периодического действия количества синтезируемого белка хватило бы примерно на 107 вакцинаций. При этом генетическая конструкция оставалась неизменной в течение 200 часов культивирования в присутствии метанола.
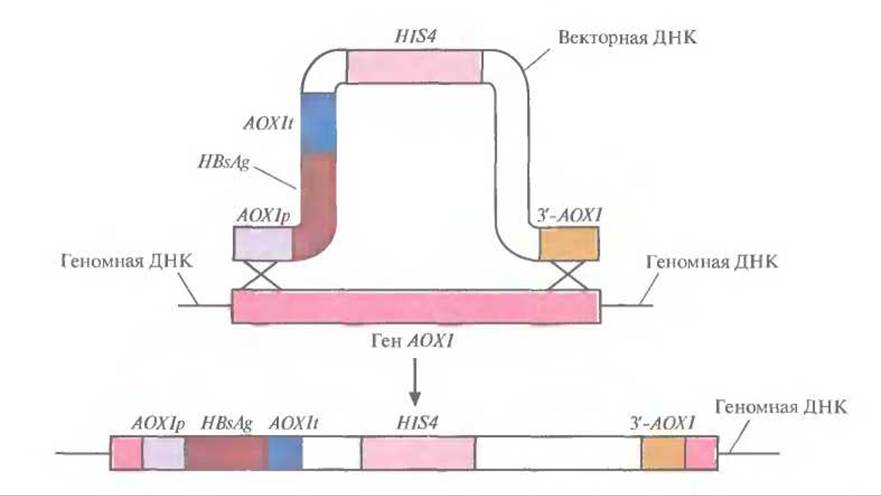
Рис. 7.6. Интеграция части экспрессирующего вектора в ген алкогольоксидазы 1 Р. pastoris. В результате двойного кроссинговера между геном АОХl и участками AOXlp и 3'-АОХ1 (верхняя часть рисунка) происходит интеграция вектора в геномную ДНК и утрата большей части гена алкогольоксидазы 1 (AОХl) хозяйской хромосомой (нижняя часть рисунка). Продукт гена HIS4 дает возможность клеткам расти на среде без гистидина. В присутствии метанола AOXlp активирует транскрипцию гена HBsAg, a AOXlt обеспечивает терминацию транскрипции и полиаденилирование.
Синтез бычьего лизоцима С2 Способность Р. pastoris секретировать гетерологичный белок исследовали в системе с использованием кДНК бычьего лизоцима С2, кодирующей полноразмерный белок и его собственный лидерный пептид. Бычий лизоцим — это желудочный фермент, разрушающий клеточные стенки бактерий; он устойчив к протеазам и сохраняет активность в узком диапазоне pH, что позволяет использовать его в качестве добавки к кормам жвачных животных для улучшения пищеварения.
Вектор, созданный для этого исследования, был идентичен вектору AOXlp-HBsAg-AOXlt, описанному выше, за исключением того, что вместо кодирующей последовательности HBsAg в него была встроена кДНК лизоцима. Вся плазмида была интегрирована в дефектную копию гена HIS4 в хромосоме Р. pastoris. В результате интеграции ген бычьего лизоцима оказывался фланкирован одним активным (HIS4) и одним дефектным (HIS4-) генами гистидинолдегидрогеназы (рис. 7.7). Предшественник бычьего лизоцима процессировался в Р. pastoris и секретировался в среду, при этом удельная активность секретируемого белка была такой же, как у нативного фермента. При ферментации 10 л культуры в течение 200 ч в непрерывном режиме при высокой плотности клеток синтезировалось примерно 20 г лизоцима.
Аутентичные гетерологичные белки были получены и с помощью других дрожжевых систем. Например, кДНК а- и ß-цепей гемоглобина А человека были встроены между промотором (МОХр) и сигналом терминации транскрипции (MOXt) гена метанолоксидазы Hansenula polymorpha и помещены друг за другом в экспрессирующий вектор. Через 40 генераций был взят изолят со случайно интегрировавшим участком исходного вектора и показано, что в нем присутствует функциональный гемоглобин А с правильной тетрамерной структурой: две а- и две ß-цепи (a2ß2). Кроме того, с использованием экспрессирующего вектора для S. pombe, несущего селективный маркерный ген и клонированный ген человека, оба под контролем промоторов млекопитающих, были получены большие количества рекомбинантных белков, кодируемых разными генами человека.
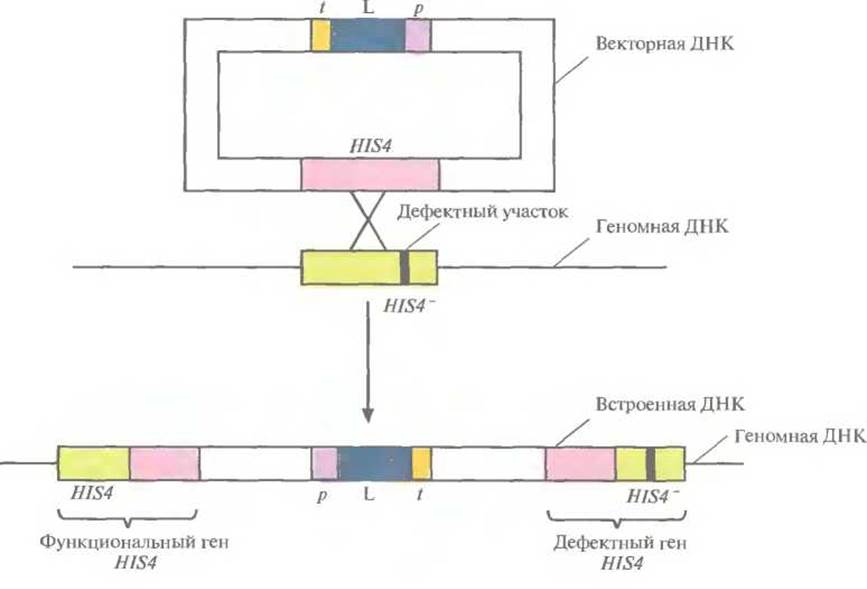
Рис. 7.7. Интеграция экспрессирующего плазмидного вектора в дефектный хромосомный ген HIS4- Р. pastoris. В результате кроссинговера между плазмидным геном HIS4 и геном HIS4-клетки-хозяина происходит интеграция в геном всей плазмиды, которая оказывается фланкированной функциональным и дефектным генами HIS4. р, L и t — промотор АОХl, кДНК бычьего лизоцима С2 и сигнал терминации транскрипции-полиаденилирования соответственно. Черная полоска — дефектный участок в HIS4-гене.
Дрожжевые системы экспрессии стали играть важную роль в получении гетерологичных белков для научных, промышленных и медицинских целей. Однако, как показали исследования, ни одна из них не может гарантировать получение аутентичного белка любого гена. По этой и ряду других причин были разработаны системы экспрессии генов с использованием клеток насекомых и млекопитающих.