Молекулярная биотехнология. Принципы и применение - Глик Б., Пастернак Дж. 2002
Молекулярная биотехнология микробиологических систем
Микробиологическое производство лекарственных средств
Ферменты
ДНКаза I
Наиболее частым летальным наследственным заболеванием среди европеоидов является муковисцидоз. В США выявлено 30 000 случаев этого заболевания, в Канаде и странах Европы — 23 000. Пациенты с муковисцидозом часто страдают инфекционными заболеваниями, поражающими легкие. Лечение рецидивирующих инфекций антибиотиками в конце концов приводит к появлению резистентных штаммов патогенных бактерий. Бактерии и продукты их лизиса вызывают накопление в легких вязкой слизи, затрудняющей дыхание. Одним из компонентов слизи является высокомолекулярная ДНК, которая высвобождается из бактериальных клеток при лизисе. Ученые из биотехнологической компании Genentech (США) выделили и экспрессировали ген ДНКазы — фермента, который расщепляет высокомолекулярную ДНК на более короткие фрагменты. Очищенный фермент вводят в составе аэрозоля в легкие больных муковисцидозом, он расщепляет ДНК, вязкость слизи снижается, что облегчает дыхание. Хотя эти меры и не излечивают муковисцидоз, они облегчают состояние больного. Применение данного фермента было недавно одобрено Департаментом по контролю за качеством пищевых продуктов, медикаментов и косметических средств (США), и объем его продаж составил в 2000 г. примерно 100 млн. долларов.
Альгинат-лиаза
Альгинат — это полисахарид, синтезируемый целым рядом морских водорослей, а также почвенными и морскими бактериями. Его мономерными единицами являются два сахарида — ß-D-маннуронат и a-L-гулуронат, относительное содержание и распределение которых и определяют свойства конкретного альгината. Так, остатки a-L-гулуроната образуют межцепочечные и внутрицепочечные сшивки путем связывания ионов кальция; остатки ß-D-маннуроната связывают ионы других металлов. Альгинат, содержащий такие сшивки, образует эластичный гель, вязкость которого прямо пропорциональна размеру полисахаридных молекул.
Выделение альгината слизистыми штаммами Pseudomonas aeruginosa существенно повышает вязкость слизи у больных муковисцидозом. Чтобы очистить дыхательные пути и облегчить состояние больных, в дополнение к обработке ДНКазой 1 следует провести деполимеризацию альгината с помощью альгинат-лиазы.
Ген альгинат-лиазы был выделен из Flavobacterium sp., грамотрицательной почвенной бактерии, активно вырабатывающей этот фермент. На основе Е. coli был создан банк клонов Flavobacterium и проведен скрининг тех из них, которые синтезируют альгинат-лиазу, путем высевания всех клонов на твердую среду, содержащую альгинат, с добавлением ионов кальция. В таких условиях весь альгинат, находящийся в среде, за исключением того, который окружает продуцирующие альгинат-лиазу колонии, образует сшивки и становится мутным. Гидролизованный альгинат теряет способность к формированию сшивок, поэтому среда вокруг синтезирующих альгинат-лиазу колоний остается прозрачной Анализ клонированного фрагмента ДНК, присутствующего в одной из положительных колоний, показал наличие открытой рамки считывания, кодирующей полипептид мол. массой около 69 000. Более детальные биохимические и генетические исследования показали, что этот полипептид, по-видимому, является предшественником трех альгинат-лиаз, вырабатываемых Flavobacterium sp. (рис. 10.3). Сначала какой-то протеолитический фермент отрезает от него N-концевой пептид массой около 6000. Оставшийся белок мол. массой 63 000 способен деполимеризовать альгинат, вырабатываемый как бактериями, так и морскими водорослями. При его последующем разрезании образуется продукт мол. массой 23 000, деполимеризующий альгинат морских водорослей, и фермент мол. массой 40 000, разрушающий альгинат бактерий. Для получения больших количеств фермента мол. массой 40 000 кодирующую его ДНК амплифицировали методом полимеразной цепной реакции (ПЦР), а затем встраивали в выделенный из В. subtilis плазмидный вектор, несущий ген, кодирующий сигнальный пептид a-амилазы В. subtilis. Транскрипцию контролировали при помощи системы экспрессии гена пенициллиназы (рис. 10.4). При трансформации клеток В. subtilis полученной плазмидой и высевании их на содержащую альгинат твердую среду с добавлением ионов кальция образовались колонии с большим ореолом. Когда такие колонии выращивали в жидкой среде, рекомбинантная альгинат-лиаза выделялась в культуральную среду. Последующие тесты показали, что этот фермент способен эффективно разжижать альгинаты, синтезируемые слизистыми штаммами Р. aeruginosa, которые были выделены из легких больных муковисцидозом. Для того чтобы определить, целесообразно ли проводить клиническое тестирование рекомбинантной альгинат-лиазы, нужны дополнительные исследования.

Рис. 10.3. Процессинг белка — предшественника рекомбинантной альгинат-лиазы Flavobacterium, происходящий в Е. coli. В результате отщепления от белка мол. массой 69 кДа пептида 6 кДа образуется белок мол. массой 63 кДа, способный деполимеризовать альгинат морских водорослей и бактериальный альгинат. Расщепление белка 63 кДа дает белок мол. массой 23 кДа, активно деполимеризующий альгинат морских водорослей, и белок мол. массой 40 кДа, гидролизующий бактериальный альгинат.
Моноклональные антитела как лекарственные средства
Примерно 100 лет назад была предпринята попытка лечения детей, больных дифтерией, с помощью неочищенной антисыворотки, полученной от лошадей, которых инфицировали Corynebacterium diphtheriae, вызывающей дифтерию у человека. С. diphtheriae инфицирует горло и миндалины, выделяя экзотоксин, приводящий к гибели клеток человека. Проникая в кровоток, этот токсин поражает органы, удаленные от места первичной инфекции, и в отсутствие лечения болезнь может иметь летальный исход. (В те времена, о которых идет речь, смертность достигала 45%.) Однако, если больному в первые несколько дней после начала инфекции ввести лошадиную антисыворотку, содержащую антитела к этому экзотоксину, то у него возникнет пассивный иммунитет, который позволяет избежать летального исхода.

Рис. 10.4. ДНК, кодирующая альгинат-лиазу мол. массой 40 кДа. К последовательности, кодирующей N-конец алыинатлиазы, присоединен сегмент гена а-амилазы В. subtilis, кодирующий ее сигнальный пептид. Транскрипция контролируется при помощи системы экспрессии гена пенициллиназы В. subtilis.
К сожалению, риск, связанный с использованием антител, не позволяет широко применять этот метод терапии. Дело в том, что в организме больного часто вырабатываются собственные антитела на чужеродные белки, присутствующие в цельной или частично очищенной антисыворотке, и ее повторное введение в случае сенсибилизации организма может привести к развитию анафилактического шока и гибели пациента.
С развитием гибридомной технологии вновь появилась надежда на то, что антитела можно будет использовать в качестве терапевтических средств для поддержания постоянного уровня чистых моноспецифичных антител в организме. Однако остаются проблемы, связанные с риском развития перекрестных реакций, приводящих к развитию иммунного ответа и анафилаксии: ведь в организме больного могут вырабатываться собственные антитела на детерминанты моноклональных антител мыши. Поэтому основная задача в настоящее время состоит в том, чтобы разработать методы получения моноклональных антител человека, обладающих как специфическими иммунотерапевтическими свойствами, так и пониженной иммуногенностью. Структура и функции антител Молекула антитела (иммуноглобулин) состоит из двух «легких» (L) и двух «тяжелых» (Н) белковых цепей, которые соединены водородными связями и расположенными в строго определенных местах дисульфидными мостиками. N-концевые участки L- и Н-цепей образуют антигенсвязывающий сайт. Отдельные домены (области) молекулы антитела выполняют разные функции, что облегчает манипуляции с генами антител (рис. 10.5). Антигенсвязывающие сайты состоят из трех участков, определяющих комплементарность антител к антигену (CDR, от англ. complementarity-determining regions), и образующих вариабельные (VH и VL) области на N-концах Н- и L-цепей. Для CDR характерна очень высокая изменчивость последовательности аминокислот, поэтому их еще называют гипервариабельными. Помимо вариабельных (VH и VL), каждая L-цепь содержит одну константную область, или домен (CL), а каждая Н-цепь — три константных области, или домена (СН1, СН2 и СH3). При обработке антитела протеолитическим ферментом папаином образуются три фрагмента: два идентичных (Fab), каждый из которых содержит интактную L-цепь, связанную дисульфидным мостиком с VH- и СНl -доменами Н-цепи, и один Fc, состоящий из двух соединенных дисульфидной связью СH2- и СH3-доменов Н-цeпи. Fab-фрагмент, точнее его N-концевая часть, называемая Fv-фрагментом, обладает антиген-связывающей активностью, присущей интактной молекуле антитела (рис. 10.5). При этом его аминокислотная последовательность у разных молекул существенно различается.
После связывания антигена с интактным антителом запускаются следующие реакции иммунного ответа.
✵ Активируется система комплемента. Компоненты этой системы разрушают клеточные мембраны, активируют фагоциты и генерируют сигналы, мобилизующие другие компоненты системы иммунного ответа.
✵ В результате связывания Fc-участка антитела с Fc-рецептором эффекторной клетки запускается реакция опосредованной антителами клеточной цитотоксичности. Активированная эффекторная клетка высвобождает вещества, лизирующие чужеродную клетку, с которой связан Fab-участок молекулы антитела.
✵ После связывания Fab-участка с растворимым антигеном Fc-участок антитела может присоединяться к Fc-рецепторам фагоцитов, которые захватывают и разрушают комплекс антиген—антитело.
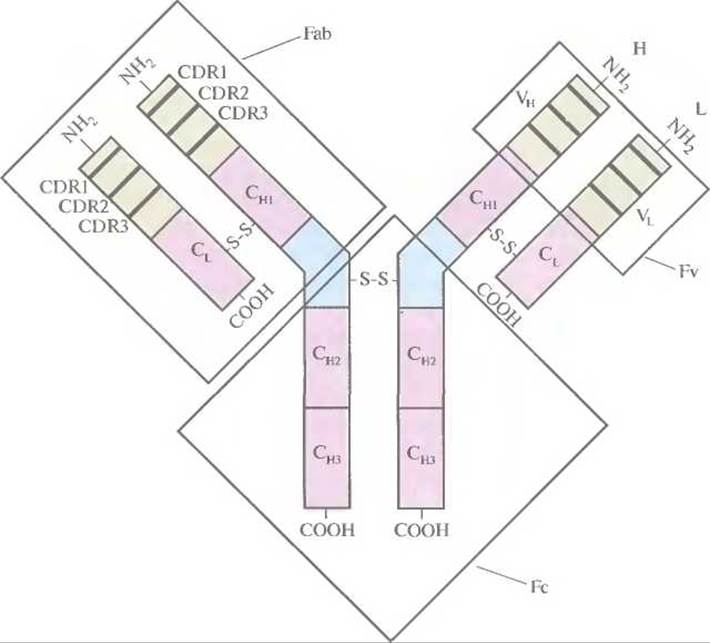
Рис. 10.5. Строение молекулы антитела. Н- и L-цепи состоят из вариабельных (VH и VL) и константных (CL, СН1, СH2 и СH3) доменов. Вариабельные домены содержат CDR-участки (CDR1, CDR2 и CDR3).
Профилактика отторжения трансплантированных органов
В 1970-х гг. были пересмотрены взгляды на пассивную иммунизацию: ее стали считать профилактическим средством борьбы с отторжением трансплантированных органов. Предлагалось вводить пациентам специфические антитела, которые будут связываться с лимфоцитами определенного типа, уменьшая иммунный ответ, направленный против пересаженного органа.
Первыми веществами, рекомендованными Департаментом по контролю за качеством пищевых продуктов, медикаментов и косметических средств (США) для использования в качестве иммуносупрессоров при пересадке органов у человека, были моноклональные антитела мыши ОКТЗ. За отторжение органов отвечают так называемые Т-клетки — лимфоциты, дифференцирующиеся в тимусе. ОКТЗ связываются с рецептором, находящимся на поверхности любой Т-клетки, который называется CD3. Это предупреждает развитие полного иммунного ответа и отторжение трансплантированного органа. Подобная иммуносупрессия весьма эффективна, хотя и оказывает некоторые побочные действия, например вызывает лихорадку и приводит к появлению сыпи.
Лекарственные вещества, связанные с моноклональными антителами
Лекарственные вещества, проявляющие высокую активность при тестировании in vitro (обычно в культуре клеток), зачастую оказываются значительно менее эффективными in vivo. Кажущееся снижение их активности объясняется тем, что они не достигают органа или клетки- мишени в нужной концентрации. Увеличение дозы принимаемого препарата не решает проблему, поскольку при этом часто возникают побочные эффекты. Более того, чтобы избежать таких эффектов, многие терапевтические средства заведомо вводят в дозах, не достигающих оптимальных, что дополнительно снижает их эффективность. Для облегчения доставки лекарственного вещества к месту его действия используют несколько приемов. 1. Заключают его в особые частицы — липосомы, липидная оболочка которых имеет высокое сродство к нужным органам. 2. Встраивают гены специфических токсинов в инфильтрирующие опухоль лимфоциты, которые высвобождают эти токсины непосредственно в опухоли. 3. Присоединяют молекулы лекарственных веществ к моноклональным антителам, специфичным по отношению к белкам, находящимся на поверхности строго определенных клеток, например опухолевых (рис. 10.6). 4. Используют лекарственные вещества в неактивной форме, переводя их в активное состояние при помощи ферментов. Чтобы такое превращение происходило только вблизи клетки-мишени, фермент присоединяют к моноклональному антителу, специфичному к поверхностному антигену этой клетки (рис. 10.6).
Для эффективной работы последней из описанных систем необходимо, чтобы а) моноклональное антитело, связанное с ферментом, переводящим лекарственное вещество в активную форму, было в достаточной степени очищено и имелось в нужном количестве; б) связывалось с высокоспецифичным для клетки-мишени белком; в) было стабильным в физиологических условиях, но в то же время быстро выводилось из кровотока; 2) при необходимости могло проникать в опухолевую ткань, обеспечивая действие препарата на все ее клетки. В этом случае мишенями оказываются строго определенные клетки, что позволяет использовать лекарственное вещество в гораздо меньших дозах, чем при прямом введении. Применение в такой системе моноклональных антител мыши может приводить к развитию иммунного ответа, поэтому очень важно использовать фрагменты антител человека или антител, максимально сходных с ними по структуре. Наиболее частой причиной смерти в странах Северной Америки и Европы является тромбоэмболия мозговых или сердечных артерий. Тромб состоит из молекул фибрина, фактора свертывающей системы крови, образующего сеть в ответ на повреждение сосудистой стенки. В норме молекулы фибрина в образовавшемся тромбе расщепляются с помощью плазмина сериновой протеиназы, который образуется из плазминогена под действием активатора (рис. 10.7). Однако нередко эта биологическая система работает недостаточно эффективно, что приводит к закупорке артерий. В таких ситуациях для повышения уровня плазмина в крови было предложено использовать активатор плазминогена в качестве терапевтического средства.
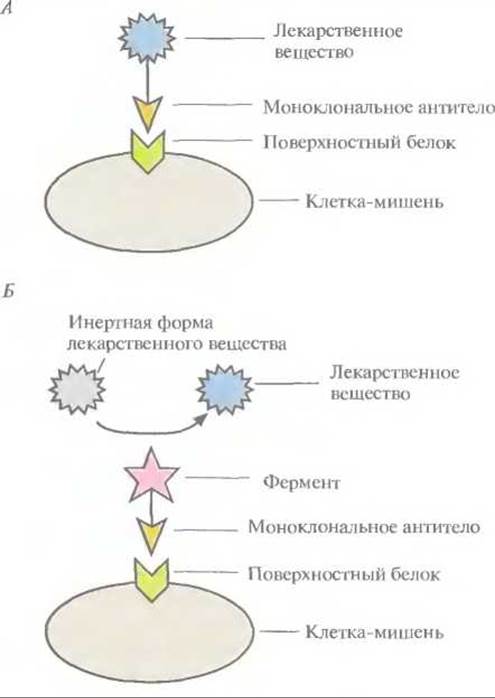
Рис. 10.6. Схематическое изображение системы целевой доставки лекарственного вещества, основанной на использовании моноклональных антител. А. Молекула лекарственного вещества присоединена к моноклональному антителу. Б. К моноклональному антителу присоединен фермент, превращающий инертную форму лекарственного вещества в активную только в непосредственной близости от клетки- мишени. В обоих случаях моноклональное антитело связывается с одним специфическим белком на поверхности клетки-мишени.
Однако плазмин способен разрушать и предшественник фибрина фибриноген (рис. 10.7), и если уровень последнего в результате терапии с использованием активатора плазминогена снизится слишком сильно, могут произойти обширные внутренние кровотечения. Это привело к необходимости создания тромболитических препаратов, разрушающих только фибрин в тромбе. Ученые исходили из того, что если к активатору плазминогена «пришить» антитело, специфичное к фибрину, то будет происходить только локальное повышение концентрации плазмина вблизи тромба (рис. 10.8). Для проверки этой гипотезы тканеспецифичный активатор плазминогена был присоединен к моноклональному антителу, специфичному в отношении фибрина. Испытания на модельных системах показали, что комплекс присоединялся к сгусткам крови и лизировал их, не вызывая значительного разрушения фибриногена. Были созданы и другие типы коньюгатов антитело—активатор плазминогена, тоже приводящие к локальному образованию плазмина, разрушающего кровяные сгустки.
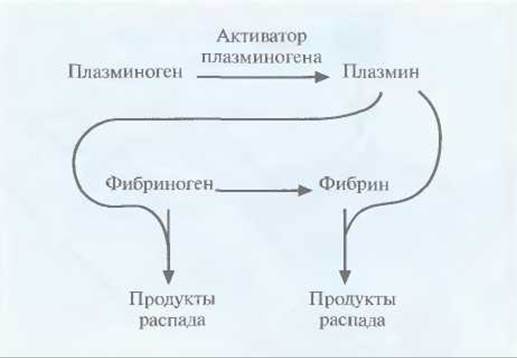
Рис. 10.7. Активация плазминогена с превращением его в плазмин и разрушение плазмином двух субстратов (фибриногена и фибрина) в крови.
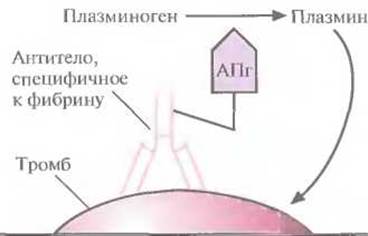
Рис. 10.8. Структура иммунотерапевтического тромболитического агента. К моноклональному антителу, специфичному к фибрину, присоединен активатор плазминогена (АПг). Этот комплекс связывается с фибрином, находящимся в тромбе, активатор плазминогена вызывает накопление плазмина вблизи тромба, и плазмин лизирует тромб.
Моноклональные антитела человека
Несмотря на кажущуюся перспективность иммунотерапии, этот метод имеет и ряд ограничений, связанных с применением моноклональных антител животных и процедурой присоединения к ним нужных молекул. Сам процесс химического присоединения весьма неэффективен, присоединение происходит случайным образом, а кроме того, при этом может снижаться ферментативная активность активатора плазминогена или других веществ, используемых в терапии. И наконец, если предполагается многократное введение препарата, необходимо использовать антитела человека, а не животных, чтобы предотвратить возникновение перекрестных иммунных реакций и сенсибилизацию пациента.
Создание специфических антител, не вызывающих перекрестных реакций, представляет собой довольно трудную задачу, поскольку получение антител человека путем традиционной гибридомной технологии сталкивается с рядом проблем.
✵ Хромосомы человека в клетках, полученных слиянием лимфоцитов человека с клетками миеломы мыши, нестабильны, поэтому трудно получить клетки, способные вырабатывать моноклональные антитела человека.
✵ Пока не удалось получить эффективые клеточные линии миеломы человека, которые могли бы заменить мышиные.
✵ Иммунизация человека различными антигенами не проводится по соображениям этического характера.
Таким образом, для получения антител человека необходимо разрабатывать другие подходы.
В одной из схем В-лимфоциты человека, активно продуцирующие специфические антитела, обработали флуоресцентно меченным антигеном, затем с помощью клеточного сортера провели обогащение образца В-лимфоцитами, вырабатывающими эти антитела. Поскольку В-клетки плохо растут в культуре, для улучшения роста их трансформировали вирусом Эпштейна—Барр. Некоторые клоны трансформированных В-клеток вырабатывают моноклональные антитела человека, взаимодействующие с селектирующим антигеном. К сожалению, выход моноклональных антител был очень небольшим и они обладали низкой антигенсвязывающей активностью. К тому же вероятность того, что в неиммунизированном организме найдутся секретирующие антитела клетки, которые будут распознавать селектирующий антиген, очень мала.

Еще один подход заключается во введении иммунных клеток человека мутантным мышам, которые практически лишены собственной иммунной системы. После трансплантации иммунных стволовых клеток человека таким мышам, страдающим тяжелым сочетанным иммунодефицитом (scid-мыши), они приобретают клетки иммунной системы человека и в ответ на введение антигена могут вырабатывать антитела человека.
Предпринимаются попытки ввести зародышам мышей гены иммуноглобулинов человека с целью создания трансгенных мышей, которые в ответ на иммунизацию конкретным антигеном смогут вырабатывать иммуноглобулины человека. Чтобы получить от трансгенных животных клетки, секретирующие специфические моноклональные антитела, можно использовать стандартную гибридомную технологию, затем провести скрининг таких положительных клеточных линий и определить, какие из них вырабатывают антитела, кодируемые генами иммуноглобулинов человека. Недавно появилось сообщение о том, что уже получена трансгенная мышь, экспрессирующая нативные формы Н- и L-цепей иммуноглобулинов человека.
Трансплантация стволовых клеток иммунной системы человека scid-мышам и получение линий трансгенных мышей — весьма трудоемкие способы производства моноклональных антител человека. Поэтому ученые пытаются создать генноинженерные методы получения антител человека, которые можно использовать в качестве терапевтических средств, и эффективных бифункциональных белков, способных связываться с мишенью и разрушать ее.
Гибридные моноклональные антитела человека и мыши
Тот факт, что разные участки молекулы иммуноглобулина выполняют разные функции, позволяет модифицировать моноклональное антителомыши таким образом, что оно приобретает некоторые сегменты антитела человека, сохраняя в то же время свою исходную антигенсвязывающую специфичность. Такое гибридное антитело называют химерным. Первым участком моноклонального антитела мыши, который был заменен соответствующим участком антитела человека, был Fc-фрагмент. Выбор объяснялся тем, что Fc-фрагмент антитела мыши выполнял роль эффектора иммунного ответа у человека недостаточно хорошо; кроме того, он с большой вероятностью индуцировал образование антител в организме человека. Чтобы снизить иммуногенность и усилить эффекторные функции, провели замену последовательностей ДНК, кодирующих Fv-области L- и Н-цепей иммуноглобулина человека, на аналогичные фрагменты специфического моноклонального антитела мыши (рис. 10.9). Такую замену можно осуществить разными путями: реплицировать ДНК in vitro с применением олигонуклеотидов в качестве затравки либо использовать субклонированные фрагменты ДНК. Сегменты ДНК, кодирующие химерные цепи, встраивали в экспрессирующий вектор и вводили в культуру В-лимфоцитов, из которой выделяли наработанные антитела.
Химерные антитела, несущие антигенсвязывающий участок моноклонального антитела мыши к поверхностному антигену клеток рака толстой кишки человека, тестировали на больных с раком толстой и прямой кишки. Антитела оставались в кровотоке примерно в шесть раз дольше обычных антител мыши, тем самым оказывая свое действие в течение большего времени. При этом лишь у одного пациента из 10 наблюдался слабо выраженный иммунный ответ. К сожалению, в этих испытаниях не удалось получить противоопухолевого эффекта антител; возможно, это было связано с введением их в слишком малых дозах или с тем, что раковый процесс находился на поздних стадиях. В опытах in vitro химерные антитела проявляли высокую эффекторную активность, что позволяет надеяться на успешное их применение в других случаях.
Конструирование химерных молекул, о которых шла речь выше, — это первый этап в создании моноклональных антител мышей и крыс, обладающих сходством с антителами человека. Другой подход состоит в замещении только CDR-участков человеческих антител фрагмента ми моноклональных антител грызунов (рис. 10.10). Такие «восстановившие форму» антитела человека могут стать эффективным терапевтическим средством, поскольку они по своей антигенсвязывающей способности приближены к исходным моноклональным антителам грызунов.

Рис. 10.9. Полученное методом генной инженерии антитело, сходное по своей структуре с антителом человека. Участки генов L- и Н-цепей иммуноглобулина человека, кодирующие VL- и VH-домены, заменены последовательностями ДНК, кодирующими VL- и VH-домены иммуноглобулина мыши. Продуктом рекомбинантных генов является химерный иммуноглобулин, обладающий антигенсвязывающей специфичностью моноклонального антитела мыши, эффекторными свойствами Fc-фрагмента иммуноглобулина человека и пониженной иммуногенностью для человека.
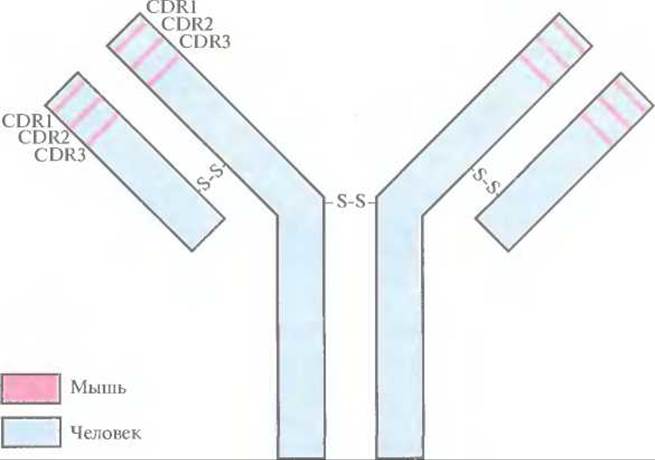
Рис. 10.10. Полученное методом генной инженерии антитело, сходное по своей структуре с антителом человека. Сегменты ДНК, кодирующие CDR-участки (CDR1, CDR2 и CDR3) антитела человека, заменены на последовательности, кодирующие CDR-участки Н- и L-цепей иммуноглобулина мыши. Продуктом этого рекомбинантного гена является иммуноглобулин с антигенсвязывающей специфичностью моноклонального антитела мыши (участки, выделенные розовым цветом) и остальными свойствами антитела человека (участки, выделенные голубым цветом).
Моноклональные антитела грызунов, сходные с антителами человека, можно получить, выделив кДНК L- и Н-цепей из клеточной линии гибридомы грызунов и амплифицировав их вариабельные области с помощью ПЦР. В качестве праймеров для амплификации можно использовать олигонуклеотиды, комплементарные высококонсервативным сегментам ДНК, фланкирующим с 5'- и 3'-концов последовательность, кодирующую вариабельную область. Зная нуклеотидные последовательности кДНК вариабельных областей легкой и тяжелой цепей (VL и VH), легко определить границы CDR, основываясь на том, что соответствующие им последовательности гипервариабельны, в то время как каркасные области относительно консервативны. Исходя из данных о нуклеотидных последовательностях ДНК, кодирующих CDR грызунов, синтезировали шесть пар олигонуклеотидных праймеров. Каждая пара инициировала синтез ДНК, кодирующей одну из шести CDR грызунов: три, локализованных на L-цепи, и три — на Н-цепи. Кроме того, на 5'-конце каждого праймера находилось 12 дополнительных нуклеотидов, комплементарных фланкирующим последовательностям каркасных участков ДНК человека, по которым происходило встраивание CDR-ДНК грызунов (рис. 10.11). Далее с помощью олигонуклеотид-направленного мутагенеза осуществляли последовательную замену CDR- ДНК человека амплифицированной CDR-ДНК грызунов — фактическую «пересадку» CDR от грызунов в каркасные участки молекулы антитела человека. Модифицированную таким образом кДНК антител встраивали в векторы экспрессии и трансформировали ими подходящие клетки-хозяева, обычно Е. coli или клетки млекопитающих, в которых и вырабатывались антитела.
Данный метод предполагает, что за антиген-связывающую способность антитела отвечают только CDR-участки, а не каркасные области. Однако, если связывание «гибридного» антитела с антигеном происходит недостаточно эффективно, может возникнуть необходимость в замене некоторых аминокислот в каркасных областях с помощью олигонуклеотид-направленного мутагенеза. К настоящему времени этим методом получено более 50 различных моноклональных антител, обладающих сходством с антителами человека. К сожалению, данная технология, являясь весьма эффективной и универсальной, довольно дорогостоящая и требует больших затрат времени. Возможно, более предпочтительным способом получения антител человека и их фрагментов окажется метод, основанный на использовании фаговых «комбинаторных» библиотек, созданных на основе мРНК, полученной из В- клеток неиммунизированных доноров.
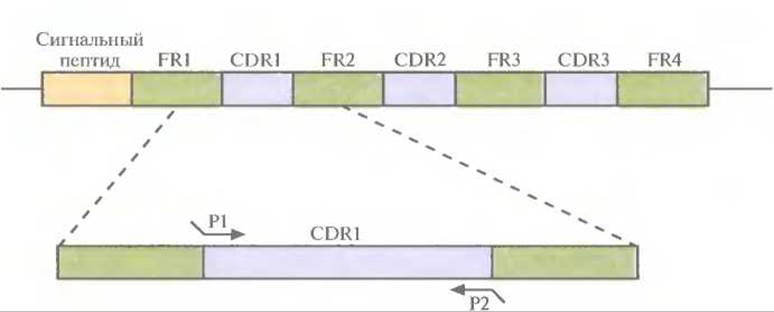
Рис. 10.11. ПЦР-амплификация CDR1-участка, входящего в состав кДНК L-цепи моноклонального антитела грызуна. Олигонуклеотидные праймеры Р1 и Р2 комплементарны последовательностям ДНК CDR1-участка антитела грызуна. Кроме того, каждый праймер содержит на 5'-конце 12 нуклеотидов, комплементарных каркасным участкам (framework region, FR) кДНК L-цепи антитела человека. Используя шесть пар олигонуклеотидных праймеров — три для VL- и три для VH-областей, — с помощью ПЦР амплифицировали отдельно каждый из ДНК-сегментов, кодирующих CDR-участки антитела грызуна. Затем, используя олигонуклеотид-направленный мутагенез, заменили ими соответствующие сегменты генов антитела человека. Такая замена оказалась возможной благодаря тому, что амплифицированные фрагменты содержали участки, комплементарные каркасным областям гена антитела человека.