Молекулярная биология клетки - Том 1 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
Молекулярная организация клеток
Плазматическая мембрана
Перенос малых молекул через мембрану
Поскольку внутренняя часть липидного бислоя гидрофобна, он представляет собой практически непроницаемый барьер для большинства полярных молекул. Благодаря такому барьеру предотвращается утечка водорастворимого содержимого клеток. Однако из-за наличия подобного барьера клетки оказались вынужденными создать специальные пути для переноса водорастворимых молекул через свои мембраны. Клетки должны получать необходимые питательные вещества и выделять вредные продукты метаболизма. Кроме того, клеткам надо регулировать внутриклеточные концентрации ионов, что подразумевает возможность транспорта определенных ионов в клетку или из клетки. Перенос малых водорастворимых молекул через липидный бислой осуществляется с помощью особых трансмембранных белков, каждый из которых отвечает за транспортировку определенной молекулы или группы родственных молекул. В клетках существуют также способы переноса через плазматические мембраны макромолекул, таких, как белки, и даже крупных частиц. Однако соответствующие механизмы сильно отличаются от механизмов транспорта малых молекул и потому будут обсуждаться в другом разделе (см. разд. 6.5).
Из данного раздела мы узнаем, что избирательная проницаемость плазматической мембраны в сочетании с активным транспортом через нее создают значительные различия в ионном составе цитозоля и внеклеточной жидкости (табл. 6-3). Это позволяет клеточным мембранам запасать потенциальную энергию в виде градиентов концентраций ионов. Трансмембранные ионные градиенты используются для осуществления различных транспортных процессов, для передачи электрических сигналов и при синтезе АТР в митохондриях, хлоропластах и бактериях. Перед тем как приступить к обсуждению свойств транспортных белков и ионных градиентов, создаваемых некоторыми из этих белков, необходимо кое-что узнать о проницаемости синтетического липидного бислоя, не содержащего белков.
Таблица 6-3. Сравнение концентраций ионов внутри и снаружи типичной животной клетки
|
Компонент |
Внутриклеточная концентрация (мМ) |
Внеклеточная концентрация (мМ) |
|
Катионы |
||
|
Na+ |
5-15 |
145 |
|
К+ |
140 |
5 |
|
Mg2+ |
0,5 |
1-2 |
|
Са2+ |
10-4 |
1-2 |
|
Н+ |
8 х 10-5 (10-7,1 М или рН 7,1) |
4 х 10-5 (10-7,4М или рН 7,4) |
|
Анионы1) Cl- |
5-15 |
110 |
1) Поскольку клетки должны содержать равное число положительных и отрицательных зарядов (чтобы быть электрически нейтральными) значительный дефицит внутриклеточных анионов отражает тот факт, что большую часть клеточного содержимого составляют отрицательно заряженные молекулы (НСО-3, РО3-, белки, нуклеиновые кислоты, метаболиты, несущие фосфатные или карбоксильные группы и т.п.). Концентрации Са2+ и Mg2+ даны для свободных ионов. В клетках имеется около 20 мМ Мg2+ и 1-2 мМ Са2+, но они в основном связаны белками и другими веществами, и в случае Са2+ хранятся внутри различных органелл.
6.4.1. Липидные бислои, не содержащие белков, непроницаемы для ионов, но свободно пропускают воду [19]
В принципе любая молекула за достаточно длительное время пройдет за счет диффузии через лишенный белков липидный бислой по градиенту концентрации. Однако скорости, с которыми различные молекулы диффундируют через такой бислой, очень сильно варьируют в зависимости главным образом от размера молекулы и ее относительной растворимости в жирах. В общем случае, чем меньше молекула и чем более она «жирорастворима» (т. е. более гидрофобна, или неполярна), тем быстрее она будет диффундировать через бислой. Малые неполярные молекулы, такие, как О2, легко растворяются в липидных бислоях и вследствие этого быстро диффундируют через них. Незаряженные полярные молекулы также диффундируют с большой скоростью, если они достаточно малы. Например, СО2 (44 Да), этанол (46 Да) и мочевина (60 Да) проходят через бислой быстро, глицерол (92 Да) - медленнее, а глюкоза (180 Да) едва ли вообще способна пройти сквозь бислой (рис. 6-41). Весьма важно то, что вода (18 Да) диффундирует через липидный бислой очень быстро, несмотря на то что молекулы воды относительно нерастворимы в жирах. Это обусловлено тем, что ее молекулы малы и незаряжены.
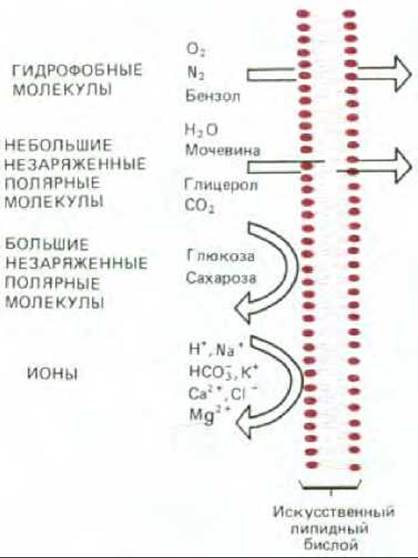
Рис. 6-41. Относительная проницаемость синтетического липидного бислоя для различных классов молекул. Чем меньше молекула и, что более важно, чем меньше она образует водородных связей, тем быстрее она диффундирует через мембрану.

Рис. 6-42. Коэффициенты проницаемости (см/с) синтетического липидного бислоя для различных молекул. Скорость потока растворенных молекул через бислой прямо пропорциональна разнице концентраций вещества на двух сторонах мембраны. Умножив разность концентраций (моль/см3) на коэффициент проницаемости (см/с), получают поток растворенного вещества в молях за секунду через квадратный сантиметр мембраны. Например, разность концентраций триптофана 10-4 моль/см3 (10-4/ 10-3 л = 0,1 М) будет обеспечивать поток 10-4 моль/см3 х 10-7 см/с = 10-11 моль/с через 1 см2 мембраны, или 6 х 104 молекул/с через 1 мкм2 мембраны.
Напротив, для всех заряженных молекул (ионов) независимо от их размеров липидные бислой оказываются в значительной степени непроницаемыми: заряд и высокая степень гидратации таких молекул препятствуют их проникновению через углеводородный участок бислоя. Вот почему искусственные бислой в 109 раз более проницаемы для воды, чем даже для таких небольших ионов, как Na+ или К+ (рис. 6-42).
6-19
6.4.2. Мембранные транспортные белки могут работать как переносчики или каналы [19]
Клеточные мембраны, так же как и искусственные липидные бислои, способны пропускать воду и неполярные молекулы за счет простой физической диффузии. Однако клеточные мембраны проницаемы и для различных полярных молекул, таких, как сахара, аминокислоты, нуклеотиды и многие другие метаболиты, которые проходят через синтетические бислои чрезвычайно медленно. За перенос подобных растворенных веществ через клеточные мембраны ответственны специфические белки, называемые мембранными транспортными белками. Они обнаруживаются во всех типах биологических мембран и могут сильно отличаться друг от друга. Каждый конкретный белок предназначен для определенного класса молекул (например, неорганических ионов, Сахаров или аминокислот), а нередко лишь какой-то разновидности молекул из этих классов. Специфичность транспортных белков была впервые показана, когда обнаружилось, что мутации в одном-единственном гене приводят к исчезновению у бактерий способности транспортировать определенные сахара через плазматическую мембрану. Аналогичные мутации теперь известны и у людей, страдающих различными наследственными болезнями, при которых нарушается транспорт тех или иных веществ в почках или кишечнике. Например, у индивидуумов с наследственной болезнью цистинурией отсутствует способность транспортировать определенные аминокислоты (включая цистин - связанный дисульфидной связью димер цистеина) из мочи или кишечника в кровь. В результате происходит накопление цистина в моче, что приводит к образованию цистиновых «камней» в почках.
Все мембранные транспортные белки, изученные настолько детально, что их расположение в мембране точно установлено, оказались трансмембранными белками, полипептидная цепь которых пересекает липидный бислой несколько раз. Эти белки обеспечивают перенос специфических веществ через мембрану без непосредственного контакта с гидрофобной внутренностью липидного бислоя, формируя в нем сквозные проходы.
Существуют два основных класса мембранных транспортных белков: белки-переносчики и каналообразующие белки. Белки- переносчики (называемые также переносчиками или транспортерами) связывают молекулу переносимого вещества, что приводит к их конформационным изменениям и как результат к переносу этой молекулы через мембрану. Напротив, каналообразующие белки (или белки-каналы) формируют заполненные водой поры, пронизывающие липидный бислой. Когда эти поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них и, следовательно, через мембрану (рис. 6-43).

Рис. 6-43. Упрощенное схематическое изображение двух классов мембранных транспортных белков. А. Белок-переносчик может попеременно существовать в двух конформациях, так что участок связывания определенного вещества последовательно доступен то с одной, то с другой стороны бислоя. Б. Каналообразующий белок формирует в липидном бислое заполненные водой поры, через которые могут диффундировать специфические ионы.
6.4.3. Активный транспорт осуществляется белками-переносчиками, связанными с источником энергии [21]
Все каналообразующие белки и многие белки-переносчики позволяют растворенным веществам проходить через мембраны только пассивно («с горки»). Этот процесс называется пассивным транспортом (или облегченной диффузией). Если молекула транспортируемого вещества не имеет заряда, то направление пассивного транспорта определяется только разностью концентраций этого вещества по обеим сторонам мембраны (градиентом концентрации). Однако если молекула заряжена, то на ее транспорт влияют как градиент концентрации, так и разница электрических потенциалов на сторонах мембраны (мембранный потенциал). Вместе концентрационный и электрический градиенты составляют электрохимический градиент. Фактически в любой плазматической мембране есть градиент электрического поля. При этом внутренняя сторона мембраны обычно заряжена отрицательно по отношению к наружной (см. разд. 6.4.15). Такой потенциал облегчает проникновение в клетку положительно заряженных ионов, но препятствует прохождению внутрь ионов, заряженных отрицательно.
Клеткам также необходимы транспортные белки, активно перекачивающие определенные растворенные вещества против их электрохимических градиентов («в горку»). Этот процесс, известный под названием активного транспорта, всегда осуществляется белками- переносчиками. Как будет рассмотрено ниже, при активном транспорте перекачивающая активность переносчиков является направленной, поскольку она тесно связана с источником метаболической энергии, таким, как гидролиз АТР или градиент ионов. Таким образом, транспорт, осуществляемый белками-переносчиками, может быть как активным, так и пассивным, в то время как транспорт через каналы является всегда пассивным (рис. 6-44).
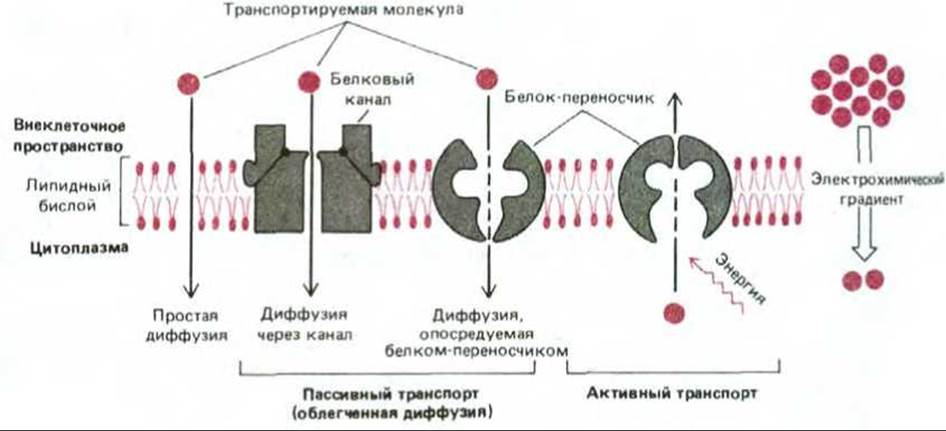
Рис. 6-44. Схематическое изображение пассивного транспорта молекул по электрохимическому градиенту и активного транспорта против. Простая диффузия и пассивный транспорт, осуществляемый транспортными белками (облегченная диффузия) протекают самопроизвольно. Для активного транспорта необходимо использовать метаболическую энергию. Только неполярные и маленькие незаряженные полярные молекулы могут проходить через липидный бислой путем простой диффузии. Перенос других полярных молекул осуществляется со значительными скоростями белками-переносчиками или каналообразующими белками.
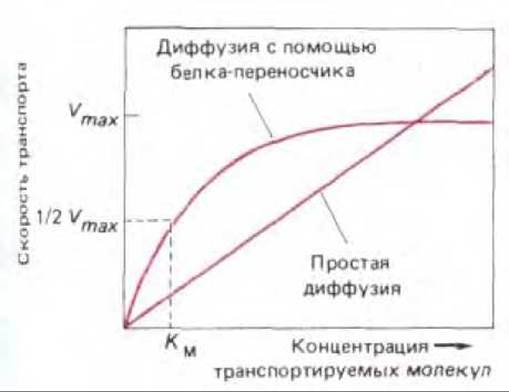
Рис. 6-45. Кинетика простой диффузии и кинетика диффузии при участии белка-переносчика. В первом случае скорость всегда пропорциональна концентрации транспортируемого вещества, а во втором скорость достигает максимального значения Vmах при насыщении белков-переносчиков. Концентрация, при которой скорость составляет половину максимального значения, принимается равной константе связывания Км молекул транспортируемого вещества данным переносчиком (аналогично Км для системы фермент-субстрат).
6-20
6.4.4. Белки-переносчики действуют как связанные с мембраной ферменты [19]
Процесс, с помощью которого белки-переносчики специфически связывают и транспортируют растворенные молекулы через липидный бислой, напоминает ферментативную реакцию, а транспортные белки выступают как особые, связанные с мембраной, ферменты. В белках-переносчиках всех типов имеются участки связывания для транспортируемой молекулы (субстрата). Когда белок насыщен (т. е. когда все участки связывания заняты), скорость транспорта максимальна. Эта скорость, обозначаемая Vmax, является характеристикой данного белка-переносчика. Кроме того, каждый белок-переносчик имеет характерную для него константу связывания Км, равную концентрации транспортируемого вещества, при которой скорость транспорта составляет половину ее максимальной величины (рис. 6-45). Связывание растворенного вещества может быть специфически блокировано как конкурентными ингибиторами (конкурирующими за тот же участок связывания), так и неконкурентными ингибиторами (связывающимися где-нибудь в другом месте и специфически влияющими на структуру переносчика). Однако в данном случае аналогия с реакцией фермент-субстрат неполная, поскольку транспортируемые вещества обычно не модифицируются ковалентно белками-переносчиками.
Некоторые транспортные белки просто переносят какое-либо растворенное вещество с одной стороны мембраны на другую. Такой простой перенос называется унипортом. Другие белки функционируют как котранспортные системы, в которых перенос одного растворенного вещества зависит от одновременного или последовательного переноса другого вещества либо в том же направлении (симпорт), либо в противоположном (антипорт) (рис. 6-46). Например, большинство животных клеток поглощают глюкозу из внеклеточной жидкости, где ее концентрация относительно высока, путем пассивного транспорта, осуществляемого специфическими переносчиками глюкозы, работающими как унипорты. Напротив, клетки кишечника и почек поглощают глюкозу из люменального пространства кишечника и почечных канальцев, где концентрация этого сахара мала. В данном случае имеет место симпорт глюкозы и ионов Na+, внеклеточная концентрация которых очень высока. Как уже обсуждалось раньше, переносчик анионов (белок полосы 3) в эритроцитах человека работает по механизму антипорта при обмене Сl- на НСО3-.
Молекулярный механизм работы белков-переносчиков пока неизвестен. Предполагается, что они переносят растворенные вещества через бислой, претерпевая обратимые конформационные изменения, которые позволяют им попеременно экспонировать участки связывания растворенных веществ то с одной, то с другой стороны. Схематическая модель того, как мог бы осуществляться этот процесс показана на рис. 6-47. Теперь мы знаем, что переносчики представляют собой трансмембранные белки, цепь которых пересекает бислой несколько раз. Маловероятно, что такие белки беспрестанно перескакивают в мембране из одного монослоя в другой или перемещаются взад-вперед через липидный бислой, как это предполагали раньше.
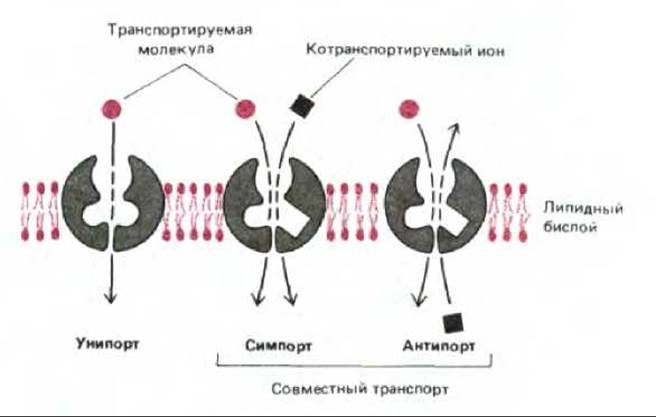
Рис. 6-46. Схема работы белков-переносчиков, функционирующих по принципу унипорта, симпорта и антипорта.
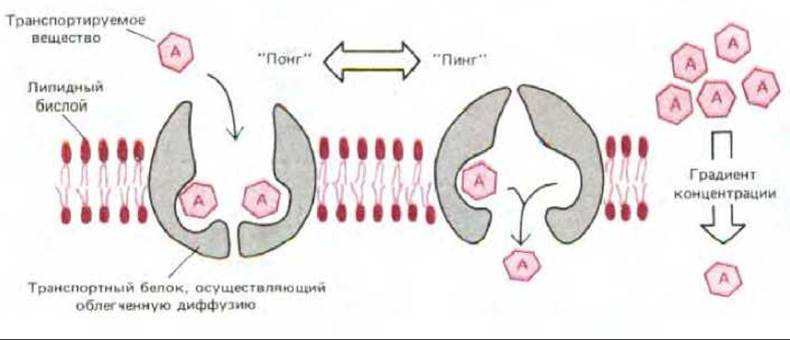
Рис. 6-47. Гипотетическая модель, показывающая как конформационные изменения в белке-переносчике могли бы обеспечить облегченную диффузию растворенного вещества А. Белок-переносчик может существовать в двух конформационных состояниях: в состоянии «понг» участки связывания для А открыты с наружной стороны бислоя; в состоянии «пинг» те же участки оказываются открытыми с другой стороны. Переход между двумя состояниями осуществляется случайным образом и полностью обратим. Поэтому при более высокой концентрации А с наружной стороны бислоя с белком-переносчиком будет связываться большее число молекул А в состоянии «понг», что приведет к транспорту вещества А по градиенту его концентрации.
Далее мы убедимся в том, что сравнительно небольшая модификация модели, изображенной на рис. 6-47, позволяет связать белок- переносчик с источником энергии, например гидролизом АТР (см. рис. 6-49). Замечательным примером белка-переносчика, использующего энергию гидролиза АТР для перекачки ионов, служит (Na+ + К+)-насос, играющий решающую роль в образовании мембранного потенциала на плазматических мембранах животных клеток.
6-23
6.4.5. (Na+ + К+)-насос плазматической мембраны - это АТРаза [22]
Концентрация К+ внутри клетки, как правило, в 10-20 раз выше, чем снаружи. Для ионов Na+ - картина прямо противоположная (см. табл. 6-3). Такая разница в концентрациях ионов обеспечивается работой (Na+ + К+)-насоса, обнаруженного в плазматических мембранах практически всех животных клеток. Этот насос работает по принципу антипорта, активно перекачивая Na+ из клеток, а К+ внутрь клеток против их крутых электрохимических градиентов. Ниже будет показано, что градиент Na+, создаваемый насосом, регулирует объем клеток за счет осмотических эффектов. Он также используется для осуществления транспорта Сахаров и аминокислот в клетку. Почти треть всей энергии, необходимой для жизнедеятельности животной клетки, тратится именно на работу этого насоса. В электрически активных нервных клетках при распространении потенциала действия происходит многократное накапливание небольших порций Na+ и потери небольших количеств К+ (см. ниже). При этом на восстановление уходит около 2/3 энергии, необходимой клетке.
Значительный шаг вперед в понимании молекулярного механизма работы натриево-калиевого насоса был сделан в 1957 г., когда обнаружилось, что для оптимальной активности фермента, гидролизующего АТР до ADP и фосфата, требуется Na+ и К+. Кроме того, было показано, что известный ингибитор (Na+ + К+)-насоса уабаин ингибирует также и АТРазу. Таким образом, была установлена связь между (Na+ + К+)-АТРазой и натриево-калиевым насосом. Однако главное доказательство того, что именно гидролиз АТР обеспечивает насос необходимой для работы энергией, было получено при изучении замкнутых теней эритроцитов. В этих опытах можно было варьировать концентрации ионов с каждой стороны мембраны и наблюдать, как эти изменения влияют на транспорт ионов и гидролиз АТР. Было обнаружено, что 1) транспорт ионов натрия и калия и гидролиз АТР тесно связаны между собой, так что ни один из этих процессов не может осуществляться без другого; 2) транспорт ионов и гидролиз АТР происходят лишь в том случае, когда N+ и АТР присутствуют внутри теней, а К+ - снаружи; 3) уабаин ингибирует АТРазу только находясь с внешней стороны теней, где он конкурирует за К+ -связывающий участок; 4) при гидролизе каждой молекулы АТР (одна молекула АТРазы может гидролизовать 100 молекул АТР за 1 с) три иона натрия выкачиваются наружу и два иона калия накачиваются внутрь (рис. 6-48).
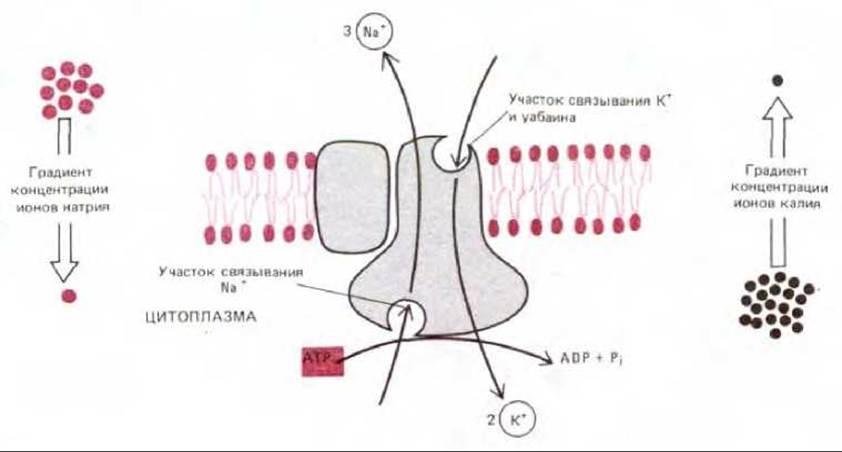
Рис. 6-48. (Na++ К+)-АТРаза активно качает Na+ наружу, а К+ внутрь клетки против их электрохимических градиентов. При гидролизе внутри клетки каждой молекулы АТР три иона Na+ выкачиваются из клетки и два иона К+ накачиваются в клетку. Специфический ингибитор насоса уабаин и ионы К+ конкурируют за общий участок на внеклеточной стороне АТРазы.
Эти эксперименты неопровержимо доказали, что АТР поставляет энергию для перекачивания ионов натрия и калия через плазматическую мембрану; тем не менее оставалось непонятным, как именно гидролиз АТР связан с транспортом ионов. Дальнейшие исследования показали, что концевая фосфатная группа АТР в присутствии Na+ переносится на остаток аспарагиновой кислоты в молекуле АТРазы. Связанная с АТРазой фосфатная группа затем гидролизуется в присутствии К+, и именно этот последний этап ингибируется уабаином. Na+-ависимое фосфорилирование сопряжено с изменением конформации АТРазы, что приводит к выведению натрия из клетки. Наоборот, К+-зависимое дефосфорилирование, осуществляемое вслед за этим, обусловливает транспорт ионов калия внутрь клетки и возвращение АТРазы в первоначальное состояние (рис. 6-49).
(Na+ + К+)-насос в тенях эритроцитов можно заставить работать в противоположном направлении - для синтеза АТР. Если градиенты концентраций ионов натрия и калия в эксперименте увеличить до такой степени, что энергия их электрохимических градиентов будет выше химической энергии гидролиза АТР, то ионы будут проходить через мембрану по их электрохимическим градиентам, а АТР будет синтезироваться из ортофосфата и ADP с помощью натриево-калиевой АТРазы. Таким образом, фосфорилированная форма АТРазы (позиция 2 на рис. 6-49) может релаксироваться либо перенося фосфат на ADP (от позиции 2 к позиции 1), либо изменяя свою конформацию (от позиции 2 к позиции 3). Будет ли общее изменение свободной энергии использоваться для синтеза АТР или же для выкачивания Na+ из теней эритроцитов, зависит от относительных концентраций АТР, ADP и фосфата и от электрохимических градиентов ионов натрия и калия.
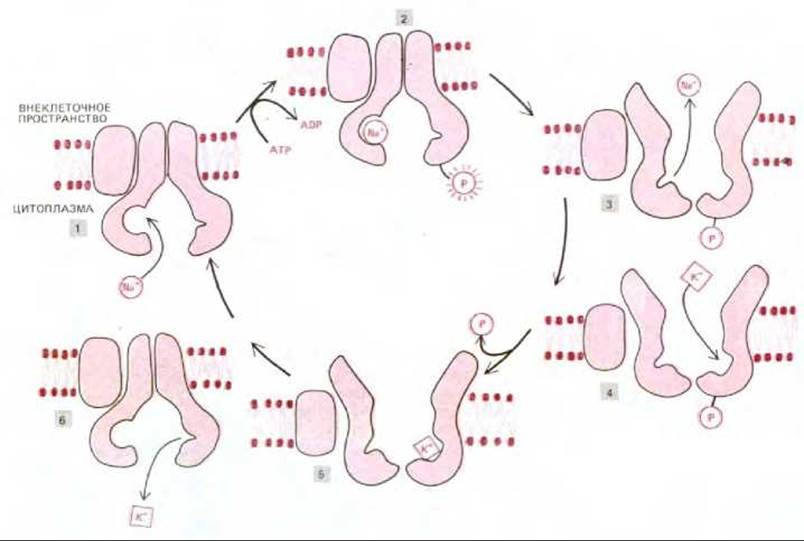
Рис. 6-49. Модель функционирования (Na+ + К+)-АТРазы. Связывание Na+ (1) и последующее фосфорилирование (2) АТРазы со стороны цитоплазмы индуцируют в белке конформационные изменения, в результате которых Na+ переносится через мембрану и высвобождается в межклеточное пространство (3). Затем связывание К+ на внешней поверхности (4) и последующее дефосфорилирование (5) возвращают белок в первоначальную конформацию; при этом К+ проходит через мембрану и высвобождается в цитоплазму (6). Эти конформационные изменения аналогичны переходам типа «пинг-понг», изображенным на рис. 6-47, за исключением того, что здесь конформационные переходы индуцируются Na+-зависимым фосфорилированием и К+-зависимым дефосфорилированием белка, вследствие чего он совершает полезную работу. Для простоты показано только по одному участку связывания Na+ и К+. В реальном насосе, видимо, существует три участка связывания Na+ и два - К+.
После того как (Na+ + К+)-АТРаза была получена в чистом виде, выяснилось, что она состоит из двух субъединиц - большой (длиной около 1000 аминокислотных остатков) трансмембранной, пересекающей бислой несколько раз и обладающей каталитической активностью, и ассоциированного с ней более мелкого гликопротеина. Первая субъединица имеет участки связывания для Na+ и АТР на цитоплазматической стороне, а для К+ и уабаина на наружной. Кроме того, она обратимо фосфорилируется и дефосфорилируется. Функция гликопротеина неизвестна. Работающий натриево-калиевый насос можно реконструировать из очищенного комплекса: АТРазу солюбилизируют в детергенте, очищают и смешивают с соответствующими фосфолипидами. После удаления детергента образуются мембранные пузырьки, которые в присутствии АТР качают Na+ и К+ в противоположных направлениях (см. рис. 6-21).
6.4.6. (Na+ + К+)-АТРаза необходима для поддержания осмотического равновесия и стабилизации объема клеток [23]
Поскольку (Na+ + К+ )-АТРаза выкачивает три положительно заряженных иона на каждые два, накачанные внутрь клетки, она оказывается «электрогенной». Это значит, что через мембрану течет ток, создающий электрический потенциал с отрицательным значением во внутренней части клетки по отношению к ее наружной поверхности. Однако этот эффект насоса дает не более 10% вклада в мембранный потенциал. Остальные 90% потенциала создаются, как мы увидим, работой насоса косвенным образом и связаны с различной концентрацией К+ по разные стороны мембраны. Повышенные концентрации калия внутри клетки необходимы для сбалансирования большого суммарного отрицательного заряда, обусловленного клеточными фиксированными анионами - множеством отрицательно заряженных органических молекул, находящихся внутри клетки и не способных проникать через плазматическую мембрану.

Рис. 6-50. Реакция эритроцитов человека на изменение осмотических условий во внеклеточной жидкости. Вода всасывается в клетку или выходит из нее по градиенту концентрации, поскольку плазматическая мембрана хорошо проницаема для молекул воды. Этот процесс называется осмосом. При помещении клеток в гипотонический раствор (т.е. в раствор с низкой концентрацией соли и, следовательно, с высокой концентрацией воды) молекулы воды движутся внутрь клеток, что приводит к их разбуханию и разрыву (лизису). Наоборот, при помещении клеток в гипертонический раствор они будут сморщиваться (см. также схему 2-1).
(Na+ + К+)-АТРаза играет непосредственную роль в регуляции клеточного объема. Она контролирует концентрацию растворов внутри клетки, а следовательно, и осмотические силы, приводящие к разбуханию или сжатию клетки (рис. 6-50). Как объясняется на схеме 6-1, растворы внутри клетки (включая фиксированные анионы и сопутствующие катионы, необходимые для уравновешивания их заряда) поддерживают большой осмотический градиент, насасывающий воду внутрь клетки. В животных клетках этот эффект нейтрализуется высокими концентрациями неорганических ионов (главным образом Na+ и Сl-), находящихся во внеклеточной жидкости. Натриево-калиевая АТРаза поддерживает осмотическое равновесие, выкачивая втекающие по ступенчатому градиенту ионы Na+ из клетки; Сl- удерживаются вне клетки благодаря мембранному потенциалу.
Важная роль (Na+ + К+)-АТРазы в регуляции клеточного объема подтверждается тем фактом, что при обработке животных клеток уабаином, ингибирующим натриево-калиевую АТРазу, они разбухают и разрываются. Осмотические проблемы могут решаться в клетках и другими способами. У многих бактерий и растительных клеток плазматическая мембрана окружена полужесткой стенкой, предохраняющей клетку от разрыва. У амеб излишек воды, проникающий внутрь в результате осмоса, собирается в сократительных вакуолях, периодически выбрасывающих свое содержимое наружу (схема 6-1). Однако в большинстве животных клеток основная роль в предотвращении разрыва из-за осмотического давления принадлежит (Na++К+)-АТРазе.
6.4.7. Некоторые Са2+-насосы - это тоже мембраносвязанные АТРазы [24]
Концентрация ионов Са2+ в цитозоле эукариотических клеток поддерживается на гораздо более низком уровне (~10-7 М) по сравнению с его концентрацией снаружи клетки (~10-3 М). Даже небольшой приток Са2+ извне значительно увеличивает концентрацию свободного Са2+ в цитозоле, а поток ионов кальция, устремляющийся по ступенчатому градиенту в ответ на внешние сигналы - один из способов передачи таких сигналов через плазматическую мембрану (см. разд. 12.3.7). Градиент Са2+ частично поддерживается с помощью существующих в плазматической мембране Са2+-насосов, активно выводящих кальций из клетки. Известно, что один из таких насосов является АТРазой, а другой работает как антипорт, обусловленный электрохимическим градиентом Na+ (см. ниже).

Схема 6-1. Внутриклеточный водный баланс: проблема и ее решение.
Наиболее изученным из Са2+-насосов является мембраносвязанная АТРаза из саркоплазматического ретикулума мышечных клеток. Саркоплазматический ретикулум образует сеть тонких каналов в цитоплазме мышечных клеток и служит внутриклеточным хранилищем ионов кальция. Когда потенциал действия деполяризует мембрану мышечной клетки, Са2+ высвобождается из саркоплазматического ретикулума в цитозоль, стимулируя мышцу к сокращению (см. разд. 11.1.4). Кальциевый насос отвечает за перекачивание Са2+ из цитозоля в саркоплазматический ретикулум. Подобно натриево-калиевому насосу, Са2+-насос - это АТРаза, которая фосфорилируется и дефосфорилируется в каждом цикле работы и накачивает два иона кальция внутрь саркоплазматического ретикулума в расчете на каждую гидролизованную молекулу АТР. Поскольку Са2+ -АТРаза - это преобладающий белок в мембране саркоплазматического ретикулума (она составляет около 90% всего белка данной структуры), ее довольно легко очистить. Оказалось, что молекула этого белка состоит из одной длинной полипептидной цепи (около 1000 аминокислотных остатков), пронизывающей мембрану несколько раз; после включения в фосфолипидные пузырьки кальциевая АТРаза осуществляет сопряженный с гидролизом АТР перенос ионов Са2+. Эксперименты по клонированию и секвенированию ДНК свидетельствуют о гомологии Са2+ -АТРазы большой каталитической субъединице (Na+ + К+)-АТРазы, что говорит об эволюционном родстве этих двух ионных насосов.
В немышечных клетках органеллы эквивалентные саркоплазматическому ретикулуму также содержат Са2+ -АТРазу, выкачивающую Са2+ из цитозоля. В ответ на специфические внеклеточные сигналы этот, изолированный от клетки, Са2+ возвращается опять в цитозоль.
6.4.8. Мембранносвязанные ферменты, синтезирующие АТР, это транспортные АТРазы, действующие в обратном направлении [25]
В плазматических мембранах бактерий, во внутренних мембранах митохондрий и тилакоидных мембранах хлоропластов обнаруживаются ферменты, очень похожие на две обсуждавшиеся выше транспортные АТРазы. Однако здесь они обычно действуют в обратном направлении. Вместо гидролиза АТР, обеспечивающего транспорт ионов, они катализируют синтез АТР из ADP и фосфата, осуществляемый благодаря наличию на этих мембранах градиента протонов. Градиент Н+ возникает на отдельных этапах транспорта электронов в процессе окислительного фосфорилирования (у аэробных бактерий и в митохондриях) или фотосинтеза (в хлоропластах), а также с помощью фотоактивируемого протонного насоса (бактериородопсина у Halobacterium). Эти ферменты, в норме синтезирующие АТР, названы АТР-синтетазами. Как и транспортные АТРазы, они способны работать в обоих направлениях в зависимости от условий: либо гидролизовать АТР и качать Н+ через мембрану во внутреннее пространство, либо синтезировать АТР при прохождении потока ионов Н+ через молекулы ферментов в обратном направлении. АТР-синтетазы ответственны за продукцию практически всего АТР в большинстве клеток и более детально обсуждаются в гл. 9.
6.4.9. Активный транспорт может осуществляться с помощью ионных градиентов [26]
Многие системы активного транспорта приводятся в действие за счет энергии, запасенной в ионных градиентах, а не путем прямого гидролиза АТР. Все ионы работают как парные транспортеры: одни функционируют по принципу симпорта, а другие - антипорта. В животных клетках котранспортируемым ионом обычно оказывается Na+. Его электрохимический градиент обеспечивает энергией активный транспорт второго вида молекул. Проникающий при этом внутрь клетки Na+ выкачивается (Na+ + К+)-АТРазой, которая, обеспечивая градиент Na+, косвенным образом осуществляет транспорт. Например, клетки кишечника и эпителиальные клетки почек содержат разнообразные симпортные системы, работающие благодаря трансмембранному градиенту Na+. Каждая система специфична для переноса внутрь клетки небольшой группы родственных Сахаров или аминокислот. В этих системах растворенные молекулы и ионы натрия связываются с различными участками на белке-переносчике; Na+ стремится войти в клетку по своему электрохимическому градиенту и как бы «тащит» молекулы сахара или аминокислоты внутрь клетки за собой. Чем выше градиент Na+, тем больше скорость всасывания растворенных молекул. Наоборот, если концентрация Na+ во внеклеточной жидкости заметно уменьшается, - транспорт растворенных молекул останавливается. Гипотетическая (и довольно упрощенная) схема функционирования подобной системы симпорта изображена на рис. 6-51.
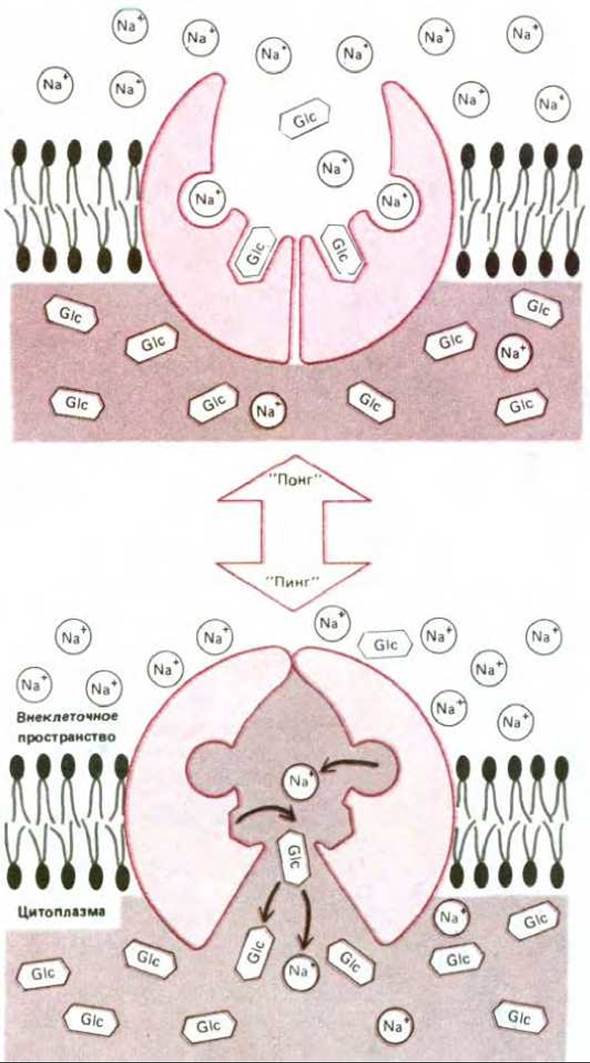
Рис. 6-51. Принципы использования градиента Na+ для работы насоса, перекачивающего глюкозу. Насос осциллирует случайным образом между двумя состояниями: «пинг» и «понг», как на рис. 6-47. Na+ связывается одинаково хорошо с белком в любой конформации. Связывание Na+ индуцирует аллостерический переход белка в состояние с сильно увеличенным сродством к глюкозе. Поскольку концентрация Na+ вне клетки выше, чем в цитозоле, связывание глюкозы с насосом более вероятно в конформации «понг». Поэтому перенос Na+ и глюкозы в клетку (переход «понг» → «пинг») происходит намного чаще, чем наоборот, т.е. осуществляется направленный перенос. Поддерживая градиент Na+ на определенном уровне (Na+ + К+)-АТРаза косвенным образом обеспечивает такую транспортную систему энергией. Говорят, что переносчики, работающие по такому принципу, осуществляют вторичный активный транспорт, тогда как АТРаза осуществляет первичный активный транспорт.
У бактерий и растений большинство систем активного транспорта, приводящихся в действие ионными градиентами, используют в качестве котранспортируемого иона Н+, а не Na+. В частности, активный транспорт большей части Сахаров и аминокислот в бактериальные клетки обусловлен градиентом Н+ через плазматическую мембрану. Наиболее хорошо изученный пример такого рода - переносчик лактозы (пермеаза). Этот трансмембранный белок, состоящий из одной полипептидной цепи (длиной около 400 аминокислотных остатков), по-видимому, пересекает липидный бислой по крайней мере девять раз. Он осуществляет Н+-зависимый симпорт: с каждой транспортируемой в клетку молекулой лактозы переносится один протон.
6.4.10. Антипорты в плазматической мембране регулируют внутриклеточное значение рН [27]
Почти все клетки позвоночных имеют в составе плазматической мембраны (Na+ + Н+)-переносчик-обменник. Он играет ключевую роль в поддержании внутриклеточного значения рН (pHi) обычно около 7,1 — 7,2. Этот переносчик обеспечивает сопряжение выброса ионов Н+ с притоком ионов Na+ и, таким образом, удаляет избыток ионов Н+, образующийся в результате клеточных реакций окисления. Работа (Na+ + Н+)- обменника регулируется значением рН: например, когда рНів мышечных клетках цыпленка выше уровня 7,7 - обменник становится неактивным; как только значение Phi падает - активность обменника увеличивается, достигая половины своей максимальной активности при рНі 7,4. Такая регуляция обусловлена связыванием Н+ с регуляторным участком обменника, находящимся на цитоплазматической стороне мембраны. Важность (Na++ К+)-обменника в поддержании уровня рНі была продемонстрирована экспериментами с мутантами фибробластов, не содержащими такого обменника: они очень быстро погибали при помещении их в достаточно кислые условия, хотя нормальные фибробласты в этих условиях жизнеспособны. В поддержании уровня рНі у многих ядерных клеток, по-видимому, играет важную роль и (Сl- + НСО3)-обменник, сходный с белком полосы 3 из мембран эритроцитов (см. разд. 6.2.6). Подобно (Na+ + Н+)-обменнику, работа (Сl- + НСО3-)-обменника регулируется значением рНі, но противоположным образом. Его активность возрастает при повышении рНі (т. е. когда цитозоль становится слишком щелочным), увеличивая скорость выведения НСО3- из клетки в обмен на Сl- и, таким образом, понижая рНі.
Существуют данные, свидетельствующие о том, что (Na+ + Н+)-обменник может участвовать не только в поддержании рНі, но и в преобразовании внеклеточных сигналов во внутриклеточные. Например, большинство белковых факторов роста в процессе стимуляции клеточной пролиферации активируют такого рода системы антипорта, увеличивая рНі от 7,1 или 7,2 до примерно 7,3. По крайней мере в некоторых случаях они делают это косвенным образом, активируя cпецифическую протеин-киназу (протеинкиназа С - см. разд. 12.3.10), которая в свою очередь фосфорилирует обменник. Это приводит к увеличению сродства регуляторного участка, связывающего Н+, и, следовательно, обменник остается активным и при больших рН. Мутантные клетки, лишенные (Na+ + Н+)-обменника, или клетки, обработанные препаратом амилоридом, ингибирующим его, не способны к ответу на действие факторов роста. Все эти факты наводят на мысль, что активация обменника с последующим увеличением рНі играет важную роль в инициации клеточной пролиферации. По-видимому, сходным образом увеличение рНі при оплодотворении яиц морского ежа, вызванное активацией (Na+ + Н+)-обменника, стимулирует синтез ДНК и белка. Остается неясным, какие внутриклеточные белки ответственны за увеличение рНі в ответ на эту активацию.
6.4.11. В основе межклеточного транспорта растворенных веществ лежит асимметричное распределение белков-переносчиков в клетках эпителия [28]
В плазматической мембране некоторых эпителиальных клеток, участвующих в поглощении пищи в кишечнике, белки-переносчики распределены асимметрично, способствуя, таким образом, сквозному транспорту поглощенных растворенных веществ через клетку. Как показано на рис. 6-52, белки, локализованные в плазматической мембране на апикальной (всасывающей) поверхности эпителиальной клетки, осуществляют Na+-зависимый симпорт, перенося питательные вещества внутрь клетки. В то же время Na+-независимые белки в базальной и латеральной мембранах позволяют питательным веществам выходить наружу по градиенту их концентраций. Градиент Na+ на плазматической мембране таких клеток поддерживается (Na+ + К+)-АТРазой, находящийся в базолатеральной области. По-видимому, сходные механизмы используются эпителиальными клетками кишечника и почек для перекачивания молекул воды из одного внеклеточного пространства в другое.
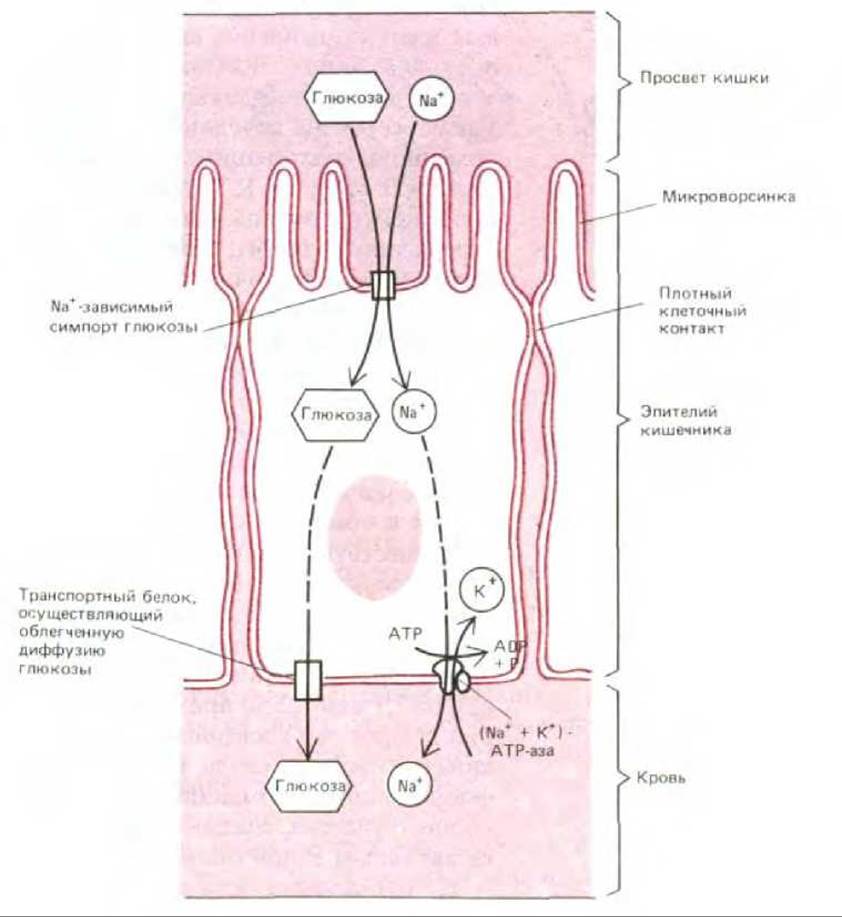
Рис. 6-52. Асимметричное распределение транспортных белков в плазматической мембране клетки эпителия кишечника приводит к сквозному транспорту глюкозы из полости кишки сквозь клетку во внеклеточную жидкость (откуда она поступает в кровь). Глюкоза проникает в клетку через апикальную мембрану посредством Na+-ависимого симпорта глюкозы и выходит из клетки (по градиенту своей концентрации) путем облегченной диффузии, опосредуемой другим белком-переносчиком глюкозы, локализованном в базальном и латеральном доменах. Градиент Na+, который приводит в движение симпорт глюкозы, поддерживается (Na+ + К+)-АТРазой, находящейся в плазматической мембране базолатерального домена; благодаря этому ферменту внутриклеточная концентрация Na+ сохраняется на низком уровне. Соседние клетки соприкасаются между собой, образуя непроницаемые контакты (называемые плотными контактами), обладающие двоякими функциями в представленном здесь процессе транспорта. Эти контакты препятствуют проникновению растворов в щели между клетками эпителия и способствуют возникновению градиента глюкозы через слой эпителиальных клеток. Плотные контакты также служат барьером внутри плазматической мембраны, препятствующим диффузии молекул по поверхности мембраны. Благодаря им различные белки-переносчики удерживаются в соответствующих мембранных доменах (см. рис. 6-36).

Рис. 6-53. Активный транспорт Сахаров внутрь бактериальных клеток за счет направленного переноса групп. Специальная «фосфотрансферазная система белков» в бактериальной мембране фосфорилирует сахар сразу после переноса его через мембрану. Донором фосфата служит фосфоенолпируват, а не АТР.
Во многих эпителиальных клетках площадь плазматической мембраны намного увеличена за счет существования тысяч микроворсинок, выступающих с апикальной поверхности в виде тонких пальцевидных образований (рис. 6-52). Такие микроворсинки могут увеличивать общую площадь всасывающей поверхности в 25 раз, тем самым значительно повышая транспортные возможности клетки. Апикальная поверхность эпителиальной клетки кишечника является также местом, где локализованы иммобилизованные гидролитические ферменты, участвующие в конечных стадиях переваривания пищи. Увеличение площади поверхности эпителия за счет микроворсинок в значительной степени способствует перевариванию и всасыванию пищи.
6.4.12. Активный транспорт в бактериях может идти путем векторного переноса групп [29]
Как мы теперь знаем, активный транспорт в клетках может приводиться в действие светом (например, в бактериородопсине), гидролизом АТР или же ионными градиентами. Четвертый путь, используемый некоторыми бактериями, состоит в том, чтобы молекулу, вошедшую в клетку посредством пассивного транспорта, «поймать в капкан» с помощью химической модификации, которая не дает возможности молекуле выйти обратно тем же путем. Например, сахара после переноса через плазматическую мембрану некоторых бактерий фосфорилируются. В результате такой модификации они становятся заряженными, не могут выйти обратно и поэтому накапливаются в клетке. Более того, вследствие фосфорилирования транспортируемых Сахаров концентрация их нефосфорилированных аналогов внутри клетки остается очень низкой, так что градиент концентрации сахара продолжает «вталкивать» эти молекулы в клетку. Поскольку фосфатные группы переносятся на молекулы растворенных веществ после их транспорта, этот тип активного транспорта называют векторным или направленным переносом групп. В большинстве хорошо изученных примеров механизм фосфорилирования достаточно сложен и тщательно регулируется. В нем принимают участие по крайней мере четыре отдельных мембранных белка, а в качестве донора высокоэнергетического фосфата используется фосфоенолпируват, а не АТР (рис. 6-53).
6.4.13. Гактерии с двойными мембранами обладают транспортными системами, которые зависят от водорастворимых субстрат-связывающих белков [30]
Как уже упоминалось выше, плазматические мембраны всех бактерий содержат белки-переносчики, использующие градиент ионов Н+ для перекачки различных питательных веществ внутрь клетки. Однако многие бактерии, включая Е. coli, имеют еще и окружающую их внешнюю мембрану, через которую растворенные вещества с молекулярной массой до 600 дальтон могут относительно свободно проникать через различные каналообразующие белки (известные под общим названием порины) (рис. 6-54). У этих бактерий для переноса некоторых Сахаров, аминокислот и мелких пептидов через внутреннюю (плазматическую) мембрану существует двухкомпонентная транспортная система, использующая водорастворимые белки, локализованные в периплазматическом пространстве между двумя мембранами. Такие периплазматические субстрат-связывающие белки присоединяют к себе специфические молекулы, которые нужно перенести через мембрану. При этом конформация белков меняется, что позволяет им связываться с другим компонентом транспортной системы - трансмембранным белком-переносчиком, находящимся во внутренней мембране (рис. 6-55). По-видимому, субстрат-связывающие белки передают присоединенное к ним растворенное вещество специальному белку-переносчику, который затем использует энергию гидролиза АТР для переноса его через внутреннюю мембрану. Те же самые периплазматические субстрат-связывающие белки служат и в качестве рецепторов в хемотаксисе -адаптивном процессе, позволяющем бактериям перемещаться в направлении более высокой концентрации специфического питательного вещества.
Теперь же мы вновь обратимся к каналообразующим белкам.

Рис. 6-54. Схематическое изображение небольшого участка двойной мембраны бактерий Е. coli. Внутренняя мембрана -это клеточная плазматическая мембрана. Между внутренней и внешней липидными бислойными мембранами находится высокопористый, упругий пептидогликан, состоящий из белков и полисахаридов, входящих в клеточные стенки бактерий. Он присоединен к липопротеиновым молекулам внешней мембраны и заполняет периплазматическое пространство. В нем содержатся различные растворимые белки. Темные нити на поверхности внешней мембраны представляют собой полисахаридные цепи специальных липополисахаридных молекул, образующих внешний монослой верхней мембраны. Для простоты показаны лишь несколько цепей. Бактерии с двойной мембраной называются грамотрицателъными, поскольку они не окрашиваются темно-синей краской по Граму. Бактерии с одной мембраной (но с толстыми клеточными стенками), например, стафиллококки или стрептококки, окрашиваются по Граму и называются грамположительными. Их единственная мембрана аналогична внутренней мембране грамотрицательных бактерий.
6.4.14. Белковые каналы образуют в плазматической мембране поры [31]
В отличие от белков-переносчиков белковые каналы (или каналообразующие белки) формируют в мембранах поры, заполненные водой. При этом каналообразующие белки внешних мембран бактерий (а также митохондрий и хлоропластов) образуют большие, относительно неспецифичные поры, а в плазматических мембранах животных и растительных клеток эти поры малы по размеру и высоко специфичны. Почти все белковые каналы служат для специфического транспорта ионов и обсуждаются здесь под названием ионных каналов. Ионные каналы обеспечивают перенос приблизительно 106 ионов в секунду, что более чем в 100 раз больше скорости транспорта, осуществляемого любым из известных белков-переносчиков. Ионные каналы никогда не работают совместно с источником энергии; осуществляемый ими транспорт всегда пассивный («с горки») и позволяет специфическим ионам, главным образом Na+, К+, Са2+ или Сl-, диффундировать по их электрохимическим градиентам через липидный бислой.
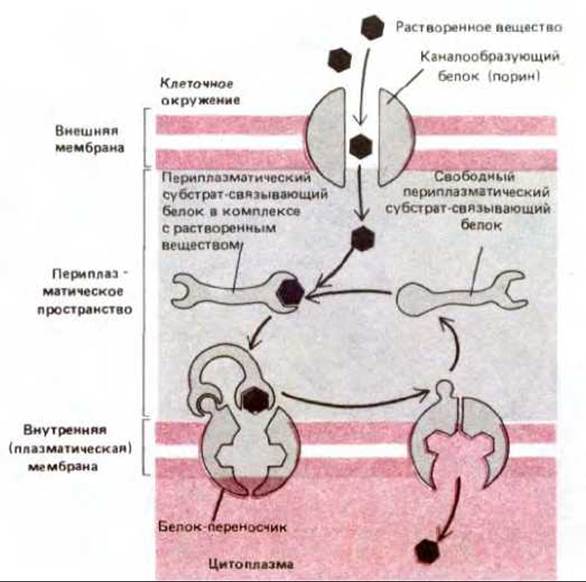
Рис. 6-55. Транспортная система, зависящая от перитазматических субстрат-связывающих белков в бактериях с двойной мембраной. Растворенные вещества диффундируют через каналообразующие белки (порины), находящиеся во внешней мембране, и связываются с периплазматическими субстрат-связывающими белками. При этом белки испытывают конформационные изменения, приобретая способность связываться с белками-переносчиками плазматической мембраны, которые затем перехватывают субстрат и активно транспортируют его через бислой. Эта стадия опосредуется гидролизом АТР. Пептидогликаны для простоты не показаны. Их пористая структура позволяет субстрат- связывающим белкам и водорастворимым веществам двигаться путем простой диффузии.
Белковые каналы плазматической мембраны обладают ионной селективностью, т. е. позволяют диффундировать через них только ионам определенного вида. По-видимому, поры должны быть достаточно узкими, чтобы ионы находились в тесном контакте с их стенками и чтобы проходить могли только те из них, которые имеют подходящий размер и заряд. Скорее всего на этом пути ионам приходится терять большинство или даже все ассоциированные с ними молекулы воды. Эти два обстоятельства накладывают ограничение на скорость диффузии через канал и делают его селективным фильтром, допускающим прохождение только ионов определенного типа. Таким образом, при увеличении концентрации ионов их поток через канал возрастает пропорционально, но лишь до определенного предела.
Другой особенностью, отличающей ионные каналы от простых пор, заполненных водой, является то, что они открыты не все время. Как показано на рис. 6-56, каналы имеют «ворота», которые открываются на короткое время, а затем закрываются. В большинстве случаев ворота открываются в ответ на специфические возмущения мембраны. Среди них наиболее известны в настоящее время изменение мембранного потенциала (потенциал-зависимые воротные каналы), механическая стимуляция (механически открываемые каналы - см. разд. 19.6.3) или связывание сигнальных молекул (лиганд-зависимые воротные каналы). Сигнальными лигандами могут быть либо внеклеточные посредники, называемые нейротрансмиттерами или нейромедиаторами (нейротрансмиттер-зависимые каналы), либо внутриклеточные посредники, например ионы (ион-зависимые каналы - см. разд. 21.1.1.), нуклеотиды (нуклеотид- зависимые каналы - см. разд. 3.3.5) или GTP-связывающие регуляторные белки (G-белок-регулируемые каналы - см. разд. 12.3.12).

Рис. 6-56. Схематическое изображение воротного ионного канала в закрытой и открытой конформациях. Трансмембранный белок, показанный в разрезе, образует в липидном бислое заполненную водой сквозную пору при открытых воротах. В состав стенок поры входят, видимо, гидрофильные аминокислотные остатки. Гидрофобные остатки взаимодействуют с липидным бислоем. Ионная селективность канала определяется самым узким его местом. Временное открытие ворот вызывается специфическим возмущением мембраны, различным для разных каналов (обсуждается в тексте). Положение ворот и ионного селективного фильтра для большинства каналов неизвестно.
На сегодняшний день известно уже около 50 видов ионных каналов. И в настоящее время продолжают открывать все новые их виды. Ионные каналы ответственны за электрическую возбудимость нервных и мышечных клеток; они осуществляют большинство форм передачи электрических сигналов в нервной системе. Отдельная нервная клетка обычно содержит более пяти видов ионных каналов. Однако такие каналы присущи не только электрически возбудимым клеткам. Они есть у всех животных клеток и обнаруживаются в некоторых клетках растений и микроорганизмов. Именно эти каналы, например, ответственны за закрывание листьев в ответ на раздражение у мимозы или за изменение направления движения на обратное у одноклеточного Paramecium.
Видимо, наиболее распространенными ионными каналами являются те, которые проницаемы главным образом для К+. Они обнаружены в плазматических мембранах почти всех животных клеток. Поскольку для их открывания скорее всего не требуется специфических мембранных возмущений, их называют иногда калиевыми проточными каналами. Эти каналы играют ключевую роль в установлении мембранного потенциала - разности электрического напряжения, наблюдающейся на двух сторонах всех типов мембран.
6-24
6-26
6.4.15. Мембранный потенциал зависит от К+-проточных каналов и градиента К+ через мембрану [32]
Мембранный потенциал зависит от распределения ионов на обеих сторонах мембраны. Выше уже шла речь о том, что (Na+ +К+)-АТРаза способствует установлению осмотического равновесия в клетке за счет поддержания низкой внутриклеточной концентрации Na+. Из-за низкой концентрации натрия внутри клетки необходим избыток других катионов, чтобы сбалансировать заряд фиксированных клеточных анионов - отрицательно заряженных органических молекул, находящихся внутри клетки. Эту роль выполняют главным образом ионы калия благодаря К+- проточным каналам, которые обеспечивают свободный переход этих ионов через мембрану и позволяют им засасываться внутрь клетки за счет отрицательного заряда фиксированных анионов (даже при отсутствии какой бы то ни было работы (Na+ + К+)-АТРазы. Таким образом, устанавливается равновесие, при котором электрическая сила, «втягивающая» ионы калия внутрь клетки, уравновешивается стремлением К+ вытекать из клетки по градиенту концентрации. Мембранный потенциал является выражением этой электрической энергии и его величина может быть рассчитана из крутизны градиента концентрации К+, необходимой для уравновешивания электрических сил. Поясним на таком примере. Предположим, что электрический градиент через плазматическую мембрану первоначально отсутствует (т. е. мембранный потенциал равен нулю), но концентрация К+ внутри клетки выше (для уравновешивания заряда фиксированных анионов), чем снаружи. В этом случае ионы калия будут стремиться выйти из клетки через проточные К+-каналы по направлению градиента их концентрации. Если ионы выйдут из клетки, то внутри останется отрицательный заряд и, таким образом, возникнет электрическое поле, другими словами, мембранный потенциал, стремящийся вернуть ионы К+ в клетку. Вытекание ионов калия прекратится, как только образовавшийся мембранный потенциал достигнет значения, при котором электрическая движущая сила, действующая на ионы калия, точно уравновесит действие градиента концентрации К+, т.е. когда электрохимический градиент ионов калия окажется равным нулю. Таким же образом создается одновременно и равновесие для ионов Сl-, но, поскольку их заряд отрицательный, они удерживаются вне клетки. Равновесные условия, при которых отсутствует электрический ток через мембрану, определяют клеточный мембранный потенциал покоя. Существует простая, но очень важная формула, количественно выражающая условия равновесия - уравнение Нернста. Как показано на схеме 6-2, она позволяет рассчитать мембранный потенциал покоя, если известно соотношение внутренней и внешней концентраций ионов.

Схема 6-2. Вывод уравнения Нернста.
Для того чтобы установился мембранный потенциал, достаточно перенести через мембрану совсем небольшое количество ионов. Таким образом, мембранный потенциал можно представить себе как перемещение зарядов, оставляющее концентрации ионов практически неизменными. В результате происходит лишь небольшое перераспределение числа положительно и отрицательно заряженных ионов между сторонами мембраны (рис. 6-57). Более того, это перемещение зарядов происходит очень быстро, за несколько миллисекунд или даже быстрее.
Рассмотрим, что случится, если инактивировать (Na+ + К+)-АТРазу. Прежде всего произойдет небольшое быстрое падение мембранного потенциала, поскольку (Na+ + К+)-насос является электрогенным и в активном состоянии вносит свой вклад в мембранный потенциал (см. разд. 6.4.6). Однако выключение этого насоса не приводит к исчезновению главного компонента потенциала покоя, основанного на механизме уравновешивания ионами калия (как описано выше). Он существует до тех пор, пока концентрация Na+ внутри клетки остается низкой, т.е. многие минуты. Но поскольку плазматическая мембрана хоть и плохо, все же проницаема для ионов Na+, то Na+ будет медленно входить внутрь клетки по своему электрохимическому градиенту. Приток натрия уменьшает мембранный потенциал и, таким образом, вызывает дополнительный отток ионов К+ из клетки. В это время нарушается осмотическое равновесие (см. разд. 6.4.6), но, если клетка не лопнула, со временем установится новое состояние равновесия между ионами Na+, К+ и Сl-. При этом мембранный потенциал будет намного ниже, чем в нормальной клетке с активным (Na+ + К+)-насосом.

Точное равенство зарядов на обеих сторонах мембраны; мембранный потенциал = 0

Несколько положительно заряженных ионов (цветные) пересекли мембрану справа налево, оставив с другой стороны отрицательно заряженные противоионы (цветные); при этом мембранный потенциал отличен от нуля
Рис. 6-57. Небольшой поток ионов несет достаточный заряд для создания большого изменения мембранного потенциала. Ионы, определяющие мембранный потенциал, располагаются вблизи мембраны, удерживаясь за счет взаимодействия с противоионами другой стороны мембраны. Для типичной клетки 1 микрокулон заряда (6 х 1012 одновалентных иона) на 1 см2 мембраны, перенесенный с одной стороны на другую, изменит мембранный потенциал примерно на I В. Это значит, что для сферической клетки диаметром 10 мкм вытекание из клетки лишь 1/100000 доли ионов К+ будет изменять потенциал на 100 мВ.
Разность потенциалов на сторонах плазматической мембраны клетки, находящейся в покое, варьирует в зависимости от организма или типа клеток от — 20 мВ до — 200 мВ. Хотя градиент К+ всегда вносит наибольший вклад в этот потенциал, значительным эффектом обладают также и градиенты других ионов (плюс неравновесные эффекты ионных насосов). Чем более проницаема мембрана для данного иона, тем в большей степени мембранный потенциал зависит от равновесных условий для этого иона. Следовательно, практически при любом изменении проницаемости мембраны для ионов происходит изменение и мембранного потенциала. Это ключевой принцип, связывающий электрическую возбудимость клеток с активностью ионных каналов.
6-25
6.4.16. Потенциал-зависимые воротные ионные каналы ответственны за электрическую возбудимость нервных и мышечных клеток [33]
В плазматических мембранах электрически возбудимых клеток (главным образом нервных и мышечных) содержится множество потенциал-зависимых воротных ионных каналов, ответственных за генерацию потенциалов действия - быстрых, скоротечных самораспространяющихся электрических возбуждений мембраны. Этот процесс начинается при деполяризации мембраны - смещении мембранного потенциала к менее отрицательному значению. Стимул, который вызывает моментальную частичную деполяризацию, сразу же открывает потенциал-зависимые воротные Na+-каналы, что позволяет небольшому количеству Na+ войти в клетку. Приток положительных зарядов в свою очередь деполяризует мембрану еще больше, приводя к открыванию других Na+-каналов, пропускающих дополнительное количество ионов натрия и, таким образом, дальнейшую деполяризацию. Этот процесс продолжается до тех пор, пока потенциал локального участка мембраны не изменится от своего значения покоя около — 70 мВ до равновесного потенциала Na+, равного примерно + 50 мВ (см. схему 6-2). При этом значении, когда суммарный электрохимический потенциал Na+ равен нулю, клетки пришли бы в новое состояние равновесия (или покоя), в котором все натриевые каналы перманентно открыты, если бы открытая конформация канала была стабильной. Однако клетки защищены от такого непрерывного электрического спазма, поскольку Na+-каналы находятся под контролем автоматического инактивирующего механизма. Они быстро закрываются после открытия, несмотря на деполяризацию мембраны. В таком инактивированном состоянии каналы не могут вновь открыться до тех пор, пока не пройдет несколько миллисекунд после падения мембранного потенциала до первоначального отрицательного значения. На рис. 6-58 схематически изображены эти три различных состояния потенциал-зависимого воротного Na+-канала - закрытое, но не инактивированное, открытое и инактивированное. Рис. 6-59 показывает, как этот канал работает при повышении и понижении потенциала действия.

Рис. 6-58. Потенциал-зависимый Na+-канал может находиться в одном из трех по крайней мере состояний (конформаций). Внутренние силы, представленные здесь в виде взаимодействия зарядов, находящихся на различных сторонах канала, стабилизируют каждое состояние и защищают от влияния небольших возмущений. Однако взаимодействия с другими молекулами могут привести к переходу из одного состояния канала в другое. Состояние с наименьшей энергией определяется мембранным потенциалом, так как различные конформаций имеют различное распределение зарядов. В состоянии покоя (мембрана сильно поляризована) канал закрыт, но не инактивирован. Это наиболее стабильное состояние с наименьшей свободной энергией. При деполяризации мембраны более низкой энергией будет обладать открытая конформация и, следовательно, канал откроется. Но свободная энергия инактивированного состояния еще ниже и после некоторого случайного периода времени в открытом состоянии канал переходит в инактивированное состояние. Таким образом, открытая конформация соответствует метастабильному состоянию, существующему недолго. Черные стрелки показывают последовательность событий при деполяризации мембраны, а красная стрелка обозначает возврат в первоначальное состояние с наименьшей свободной энергией после реполяризации мембраны.
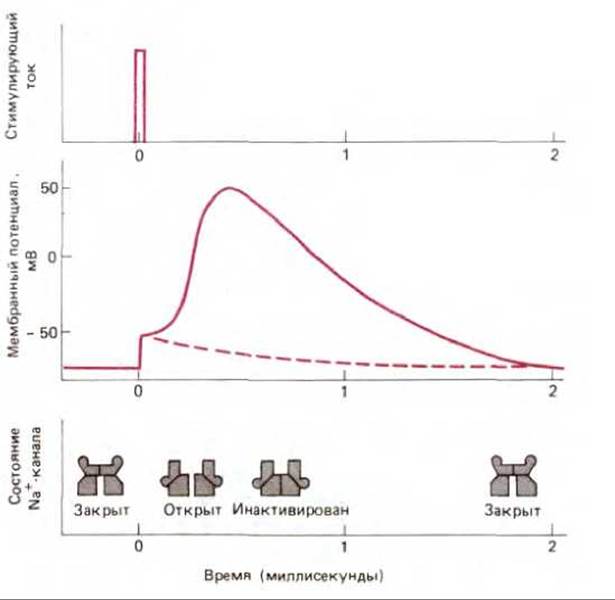
Рис. 6-59. Индукция потенциала действия коротким электрическим импульсом (показан на верхнем графике). Импульс частично деполяризует мембрану (средний график). Сплошная линия на графике мембранного потенциала показывает возникновение потенциала действия при открывании и последующей инактивации потенциал-зависимых Na+-каналов. Мембранный потенциал автоматически возвращается к своему первоначальному значению — 70 мВ при закрытии Na+-каналов благодаря непрерывному вытеканию К+ через К-каналы. Возникновение второго потенциала действия невозможно до тех пор, пока Na+-каналы (их состояния показаны внизу) не вернутся в закрытое, но не инактивированное состояние (см. рис. 6-58). До этого мембрана остается нечувствительной к раздражению. Прерывистая линия показывает релаксацию мембранного потенциала при слабых воздействиях, не приводящих к открыванию каналов.
Рассмотренный выше процесс образования потенциала действия относится лишь к небольшому участку плазматической мембраны. Однако самоусиливающейся деполяризации этого участка достаточно для деполяризации соседних областей мембраны, которые вовлекаются при этом в тот же цикл генерации потенциала действия. Таким образом, потенциал действия распространяется от первоначального участка деполяризации по всей плазматической мембране. Более детальное рассмотрение функций и свойств потенциала действия приводится в гл. 19.
В нейронах и мышечных клетках имеется несколько тысяч потенциал-зависимых Na+-каналов. Ток, протекающий через мембрану, - это сумма всех микротоков через каждый канал. Суммарный ток можно записать с помощью микроэлектрода (см. разд. 19.2.3). Однако существует возможность зарегистрировать и токи, протекающие через индивидуальные каналы. Это делают с помощью специальной техники, позволяющей выделить совсем небольшой участок мембраны, который содержит лишь несколько каналов, и записать затем протекающие через него токи. Такая методика (patch-clamp) дает более детальную картину работы этих каналов.
6-27
6.4.17. Регистрация токов, проходящих через изолированный участок мембраны, показывает, что индивидуальные Na+-каналы открываются по принципу «все или ничего» [34].
Разработка метода patch-clamp позволила существенно продвинуться в изучении ионных каналов. С помощью этого метода можно анализировать транспорт через единичную молекулу белкового канала, находящегося на маленьком участке мембраны (рис. 6-60), записывать сигналы от ионных каналов любых типов клеток, включая и электрически невозбудимые. Многие из этих клеток, например дрожжевые, слишком малы, чтобы исследовать их традиционным методом электрофизиологов - введением внутриклеточных электродов.
Регистрация сигналов методом patch-clamp показала, что индивидуальные Na+-каналы открываются по принципу «все или ничего». В открытом состоянии их проводимость не меняется, а времена открывания и закрывания случайны. Поэтому суммарный ток, протекающий через большую популяцию Na+-каналов мембраны целой клетки дает представление не о степени открытия индивидуального канала, а лишь о средней вероятности того, что он открыт (рис. 6-61).
Явление потенциал-зависимого открывания и закрывания можно понять исходя из простых физических принципов. Внутри покоящейся нервной или мышечной клетки электрический потенциал на 50- 100 мВ ниже, чем снаружи. Такая разность потенциалов на двух сторонах мембраны может показаться незначительной, однако, учитывая, что толщина мембраны составляет всего лишь около 5 нм, градиент оказывается равным примерно 100 000 В/см. Следовательно, мембранные белки находятся в очень сильном электрическом поле. Естественно, мембранные белки, как и все другие, содержат на своей поверхности некоторое количество заряженных групп. Электрическое поле увеличивает силы, действующие на структуру молекулы. На многие мембранные белки изменения электрического поля через мембрану не оказывают значительного влияния. Ионные каналы, однако, приобрели в процессе эволюции тонкую сбалансированную чувствительность к электрическому полю: они могут принимать несколько альтернативных конформаций, стабильность которых зависит от величины электрического поля. Малые возмущения не отражаются на конформации каналов, но при достаточно сильных воздействиях, например случайных тепловых движениях окружающих молекул, может произойти и переход к другой конформации (см. рис. 6-58).
Функции потенциал-зависимых Na+-каналов специфически блокируются двумя паралитическими ядами: тетродотоксином (ТТХ), получаемым из иглобрюхих рыб и сакситоксином, который выделяют из определенных видов морских динофлагеллят. Из-за высокой аффинности и специфичности эти токсины оказались незаменимыми для фармакологических исследований, подсчета числа Na+-каналов в мембране и для очистки этих каналов. Было показано, что в плазматической мембране клеток скелетных мышц находится лишь несколько сотен Na+-каналов на 1 мкм2, т.е. один канал на 10 000 молекул фосфолипидов. Несмотря на такую малую плотность каналов, эти мембраны электрически возбудимы, поскольку каждый канал обладает высокой проводимостью, пропуская более 8000 ионов за 1 миллисекунду.
В 1984 году была определена нуклеотидная последовательность ДНК, детерминирующая образование потенциал-зависимого Na+-канала (у угря). Установлено, что она кодирует одну длинную полипептидную цепь (около 1800 аминокислотных остатков), содержащую четыре гомологичных трансмембранных домена (каждый из которых имел в своем составе шесть предполагаемых а-спиралей, пронизывающих мембрану). Эти спирали, по-видимому, взаимодействуют друг с другом, образуя стенки поры, заполненной водой. Совсем недавно был секвенирован ген, кодирующий потенциал-зависимый Са2+-канал. Оказалось, что это также длинный полипептид, первичная структура которого высоко гомологична обнаруженной для Na+-канала. Весьма вероятно, что потенциал-зависимые ионные каналы относятся к семейству эволюционно и структурно родственных белков. В каждом из этих каналов один из предполагаемых трансмембранных сегментов содержит положительно заряженные аминокислотные остатки, разделенные между собой регулярными промежутками. Не исключено, что эти остатки вместе обладают функцией сенсора потенциала, обеспечивая открывание канала в ответ на достаточную деполяризацию мембраны (см. рис. 6-58).
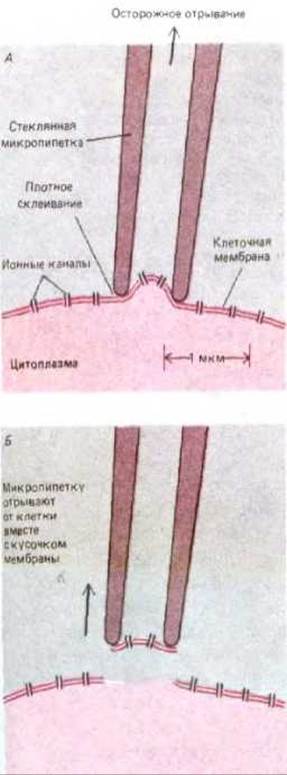
Рис. 6-60. Запись сигналов методом patch-clamp. Благодаря плотному соединению между микропипеткой и мембраной ток может течь только через каналы, имеющиеся в участке мембраны, закрывающей кончик пипетки. Запись протекающего тока можно сделать как в случае А (в интактной клетке), так и в случае Б (удалив участок мембраны). Преимущества варианта Б состоят в легкости изменения условий с любой стороны мембраны для изучения воздействия различных растворов на поведение канала. Ориентацию удаленного участка мембраны можно изменить на противоположную (см. также рис. 4-33 и 4-34).

Рис. 6-61. Запись тока, протекающего через единичный потенциал-зависимый Na+-канал, находящийся в крошечном участке плазматической мембраны мышечной клетки эмбриона крысы (см. рис. 6-60). Мембрану деполяризуют импульсом (А). Три графика тока (Б) получены в трех экспериментах с одним и тем же участком мембраны. Каждое существенное изменение тока соответствует открытию и закрытию одного канала. Сравнение показывает, что время открытию и закрытию может существенно варьировать, при этом скорость протекания зарядов через канал остается практически постоянной. Маленькие флуктуации при записи тока являются электрическим шумом записывающей аппаратуры. Суммарный ток, записанный в 144 повторяющихся экспериментах, показан на В. Он эквивалентен току Na+ через относительно большой участок мембраны, содержащий 144 канала. Сравнение Б и В показывает, что суммарный ток отражает вероятность открывания индивидуального канала. Эта вероятность со временем уменьшается, так как каналы деполяризованной мембраны переходят в инактивированную конформацию. Кинетика открывания и инактивации каналов мышечной клетки эмбриона намного медленнее, чем у типичной нервной клетки. (По данным J. Patlak и R. Horn, J. Gen. Physiol., 79, 333-351, 1982, с разрешения Rockefeller University Press.)
Намного больше известно о структуре другого класса ионных каналов, открывающихся в ответ на связывание специфических нейротрансмиттеров, а не на изменения мембранного потенциала. Эти трансмиттер-зависимые ионные каналы также принадлежат к одной группе родственных белков. Однако в отличие от потенциал-зависимых Na+ - и Са2+-каналов, каждый из которых образован одной длинной полипептидной цепью, все изученные трансмиттер-зависимые ионные каналы построены из нескольких гомологичных субъединиц.
6.4.18. Ацетилхолиновый рецептор - это трансмиттер-зависимый катионный канал [35]
Трансмиттер-зависимые ионные каналы приспособлены для превращения внеклеточных химических сигналов в электрические сигналы. Они располагаются обычно в специализированных соединениях (называемых химическими синапсами), расположенных между нервными клетками и клетками-мишенями. Эти каналы концентрируются на плазматической мембране клетки-мишени в области синапса. Каналы способны открываться на некоторое время в ответ на связывание нейротрансмиттера, высвобождаемого нервным окончанием. При этом меняется проницаемость постсинаптической мембраны клетки-мишени (рис. 6-62). В отличие от потенциал-зависимых каналов, ответственных за возникновение потенциалов действия, трансмиттер-зависимые каналы относительно нечувствительны к мембранному потенциалу и поэтому неспособны к самоусиливающемуся возбуждению. Вместо этого они изменяют проницаемость мембраны и, следовательно, влияют на мембранный потенциал. Величина этого изменения зависит от того, сколько трансмиттера высвободилось в синапсе и в течение какого времени он там присутствует. Ясно, что потенциал действия может возникнуть только при условии, что потенциал-зависимые каналы также присутствуют в этой же мембране клетки-мишени.
Кроме характерной ионной селективности каждый трансмиттер-зависимый канал обладает высокоспецифичным участком связывания своего нейротрансмиттера. Примером наиболее изученного трансмиттер-зависимого канала может служить ацетилхолиновый рецептор клеток скелетных мышц. Этот канал временно открывается при действии ацетилхолина, нейротрансмиттера высвобождаемого из нервного окончания в нервно-мышечное соединение (см. разд. 19.3.1). Ацетилхолиновый рецептор занимает особое место в истории изучения ионных каналов. Он был первым ионным каналом, выделенным в чистом виде, именно у него впервые была определена полная аминокислотная последовательность, ацетилхолиновый рецептор оказался первым каналом, для которого удалось добиться функциональной активности после реконструкции в синтетическом липидном бислое, и, наконец, первым каналом, у которого был записан электрический сигнал, получаемый при открывании одного канала. Ген этого канала также оказался первым из генов белков-каналов, которые были выделены, клонированы и секвенированы. Успех в изучении этого рецептора стал возможен по двум по крайней мере причинам. Во-первых, существует необычайно богатый источник для его выделения из электрических органов электрических рыб и скатов. Эти органы представляют собой модифицированные мышцы, приспособленные для того, чтобы вызвать у жертвы электрический шок. Во-вторых, некоторые нейротоксины типа а-бунгаротоксина из яда определенных змей с высокой эффективностью (Ка = 109 л/моль) и специфичностью связываются с этим рецептором и могут быть использованы для его очистки методом аффинной хроматографии. Благодаря применению флуоресцентно или радиоактивно меченного а-бунгаротоксина показано, что ацетилхолиновые рецепторы плотно упакованы в плазматической мембране мышечных клеток в месте нейромышечного соединения (около 20000 рецепторов на мкм2), а в других местах той же мембраны находится лишь несколько таких рецепторов.
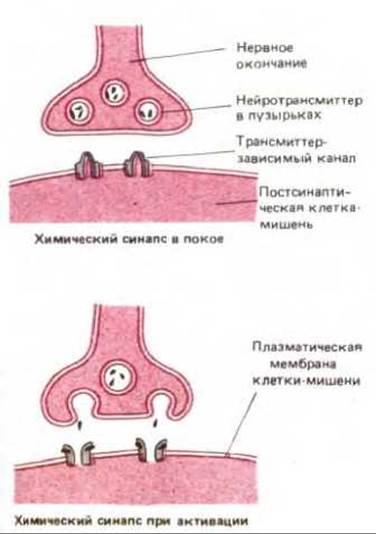
Рис. 6-62. Химический синапс. Приходящий к нервному окончанию потенциал действия стимулирует высвобождение нейротрансмиттера, содержащегося в секреторных пузырьках и высвобождаемого из клетки при слиянии пузырьков с плазматической мембраной нервного окончания. Высвобожденный нейротрансмиттер связывается с трансмиттер-зависимыми ионными каналами, сконцентрированными на плазматической мембране постсинаптической клетки, и открывает их. В результате тока ионов изменяется мембранный потенциал клетки-мишени. Таким образом происходит передача нервного сигнала.

Занят и закрыт Занят и открыт
Рис. 6-63. Три конформации ацетилхолинового рецептора. Связывание двух молекул ацетилхолина открывает ворота трансмиттер-зависимого ионного канала. Но, по-видимому, даже при связанном ацетилхолине рецептор остается открытым непродолжительное время, а затем закрывается. Ацетилхолин отсоединяется от рецептора, возвращая его в первоначальное состояние.
Ацетилхолиновый рецептор представляет собой гликопротеин, состоящий из пяти трансмембранных полипептидов. Два из них принадлежат к одному типу, а три остальных - к другому. Они кодируются четырьмя различными генами. Поскольку четыре этих гена обнаруживают тесную гомологию, предполагают, что все они произошли от одного гена-предшественника. Два идентичных полипептида в пентамере имеют участки связывания ацетилхолина. При связывании двух молекул трансмиттера с пентамерным комплексом происходит индуцированное конформационное изменение, приводящее к открыванию канала. Канал открывается примерно на 1 миллисекунду, а затем опять закрывается. По-видимому, как и для потенциал-зависимого Na+-канала, открытая форма является короткоживущей и быстро переходит в закрытое состояние с меньшей свободной энергией (рис. 6-63). После этого молекулы ацетилхолина диссоциируют из комплекса с рецептором и гидролизуются специфическим ферментом (ацетилхолинэстеразой). Освободившись от связанного нейротрансмиттера, ацетилхолиновый рецептор возвращается к исходному состоянию покоя.
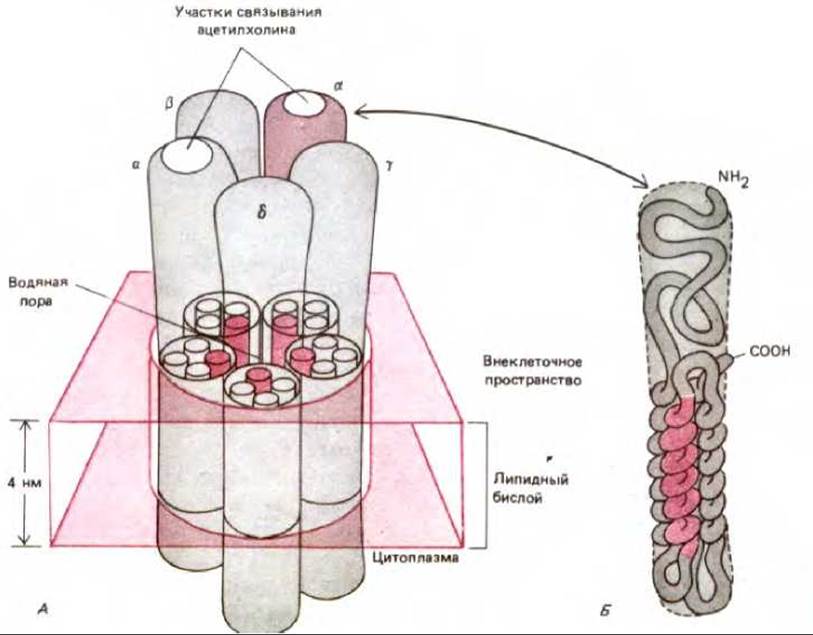
Рис. 6-64. Одна из моделей образования трансмембранной поры, заполненной водой, из пяти гомологичных субъединиц (а, а, ß, у, δ) ацетилхолинового рецептора (А). Обратите внимание, что обе а-субъединицы содержат участок связывания ацетилхолина и что основная масса рецептора находится во внеклеточном пространстве. Каждая субъединица состоит из ~500 аминокислотных остатков. Мг рецептора ~300 000 Да. Предполагается, что полипептидная цепь каждой субъединицы пересекает липидный бислой в виде четырех а-спиралей (Б). Одна из спиралей (показана в цвете) содержит более полярные аминокислотные остатки, чем другие. Она, видимо, и входит в состав стенки водяной поры при объединении пяти субъединиц (А).
Для изучения структуры ацетилхолинового рецептора были использованы методы электронной микроскопии и малоугловой дифракции рентгеновских лучей, однако точный ответ на вопрос, как образуется трансмембранный гидрофильный канал, до сих пор не получен. Было предложено несколько моделей, основанных главным образом на аминокислотной последовательности субъединиц. Одна из моделей представлена на рис. 6-64. То, что кластеры отрицательно заряженных аминокислотных остатков выстилают отверстие канала, объясняет, по-видимому, известный факт, что отрицательно заряженные ионы не способны проходить через канал, а положительно заряженные ионы с размером до 0,65 нм могут это делать. Через канал проходят преимущественно ионы Na+ и К+, а также некоторое количество Са2+. Строгих ограничений на вид катионов не существует, поэтому поток каждого из них через канал определяется главным образом их концентрациями и электрохимическими движущими силами. Так как градиент напряжения уравновешивает градиент концентрации К+ через мембрану при наличии потенциала покоя, то и движущая сила для ионов К+ близка к нулю (см. схему 6-2). Напротив, для ионов Na+ как градиент напряжения, так и градиент концентрации действуют в одном направлении, способствуя движению ионов внутрь клетки. Это же справедливо и для Са2+, но его внеклеточная концентрация намного меньше концентрации ионов натрия, и, следовательно, вклад Са2+ в общий ток ионов незначителен. Поэтому открывание ацетилхолиновых рецепторных каналов приводит к большому притоку ионов Na2+ (максимальная скорость притока составляет около 30 000 ионов на 1 канал за 1 миллисекунду). Этот ток вызывает деполяризацию мембраны, что служит сигналом для мышечного сокращения, как описано ниже.
Ранее были определены также последовательности нуклеотидов ДНК, кодирующие субъединицы нескольких разны трансмиттер-зависимых ионных каналов. Выведенные из них аминокислотные последовательности гомологичны друг другу и соответствующим субъединицам ацетилхолинового рецептора, что говорит об эволюционном родстве этих ионных каналов.
6.4.19. Нервно-мышечная передача включает в себя последовательную активацию по крайней мере четырех различных наборов воротных каналов [36]
Исключительную роль ионных каналов, имеющих «ворота» (или воротных каналов) для работы электрически возбудимых клеток, можно проиллюстрировать на примере стимуляции мышечной клетки к сокращению приходящим нервным импульсом. Этот с виду простой ответ состоит из последовательного открывания и закрывания по крайней мере четырех различных наборов каналов, имеющих ворота, и все это происходит менее чем за 1 секунду (рис. 6-65).
1. Процесс начинается, когда нервный импульс достигает нервного окончания и деполяризует его плазматическую мембрану. Деполяризация открывает на время потенциал-зависимые воротные Са2+-каналы в этой мембране. Поскольку концентрация Са2+ снаружи клетки более чем в 1000 раз превышает концентрацию свободного Са2+ в клетке, ионы кальция устремляются внутрь нервного окончания. Увеличение концентрации Са2+ в цитозоле нервного окончания стимулирует локальное высвобождение ацетилхолина в синаптическую щель.
2. Высвобожденный ацетилхолин связывается с ацетилхолиновыми рецепторами на плазматической мембране постсинаптической мышечной клетки. Это вызывает временное открывание катионных каналов рецепторов. В результате приток Na+ приводит к локальной деполяризации мембраны мышечной клетки.
3. Деполяризация плазматической мембраны мышечной клетки открывает ворота потенциал-зависимых Na+-каналов этой мембраны, способствуя засасыванию еще большего количества ионов Na+. Таким образом происходит усиление деполяризации мембраны. Это в свою очередь приводит к тому, что открываются следующие потенциал-зависимые Na+-каналы и в конце концов возникает волна деполяризации (потенциал действия), которая распространяется до тех пор, пока не охватит всю мышечную мембрану.
4. Общая деполяризация плазматической мембраны мышечной клетки приводит к временному открытию Са2+-каналов в мембранах саркоплазматического ретикулума и высвобождению Са2+ в цитозоль. В результате происходит повышение внутриклеточной концентрации Са2+, вызывающее сокращение миофибрилл в мышечной клетке (см. разд. 11.1.11). Пока неизвестно, каким образом изменения напряжения на мышечной плазматической мембране служат сигналом для открывания потенциал-зависимых Са2+-каналов в мембране саркоплазматического ретикулума. Другая возможность связана с тем, что деполяризация мышечной плазматической мембраны активирует медиаторные пути передачи сигнала с помощью инозитолфосфолипида, что обсуждается в гл. 12.
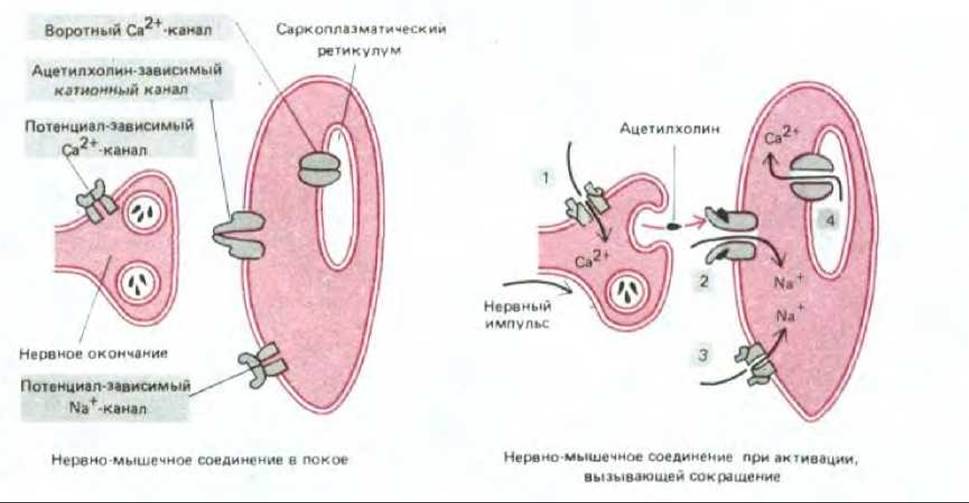
Рис. 6-65. Схема нервно-мышечного соединения, показывающая, как некоторые имеющие ворота каналы участвуют в стимуляции мышечного сокращения нервным импульсом. Каналы пронумерованы в той последовательности, в которой они открываются (см. текст). Механизм открывания Са2+-каналов в саркоплазматическом ретикулуме неизвестен.
6.4.20. Ионофоры повышают ионную проницаемость мембран [37]
Ионофоры - это небольшие гидрофобные молекулы, которые растворяются в липидных бислоях и повышают их проницаемость для ионов. Большинство ионофоров синтезируется микроорганизмами (вероятно, в качестве оружия против своих конкурентов), некоторые из них используются как антибиотики. Ионофоры широко применяются в клеточной биологии для повышения проницаемости мембран по отношению к определенным ионам в исследованиях на синтетических бислоях, клетках и клеточных органеллах. Существуют два класса ионофоров - подвижные переносчики ионов и каналообразующие ионофоры (рис. 6-66). Ионофоры обоих типов действуют, экранируя заряд транспортируемого иона так, чтобы последний мог пройти гидрофобную внутреннюю область липидного бислоя. Поскольку ионофоры не связаны ни с какими источниками энергии, они лишь позволяют ионам двигаться по их электрохимическим градиентам.
Примером подвижного переносчика ионов может служить валиномицин. Он представляет собой полимер, повышающий проницаемость мембраны для ионов К+. Валиномицин имеет кольцеобразную структуру. Наружная гидрофобная часть его молекулы состоит из боковых цепей валина и контактирует с углеводородной сердцевиной липидного бислоя. Во внутренней полярной области как раз может поместиться один ион калия (рис. 6-67). Валиномицин переносит К+ по его электрохимическому градиенту, он захватывает этот ион с одной стороны мембраны, диффундирует с ним через бислой и высвобождает его на другой стороне.
Еще один пример подвижного переносчика ионов-ионофор А23187, который транспортирует двухвалентные катионы, такие, как Са2+ и Mg2+. Этот ионофор обычно действует как ионообменный «челнок»: на каждый двухвалентный катион, вносимый им в клетку, он удаляет два иона Н+ из клетки. Если клетки подвергнуть действию ионофора А23187, ионы Са2+ устремляются в цитозоль по крутому электрохимическому градиенту. Поэтому ионофор А23187 широко используют в клеточной биологии для повышения концентрации свободного Са2+ в цитозоле, моделируя, таким образом, определенные медиаторные механизмы передачи сигнала в клетке (см. разд. 12.3.10).
Если температура мембраны опускается ниже точки ее замерзания, подвижные переносчики уже не могут диффундировать через липидный бислой, и ионный транспорт прекращается. Наличие такой температурной зависимости свидетельствует о том, что данный ионофор - это подвижный переносчик. Если же транспорт ионов продолжается даже в замороженном бислое, можно сделать вывод, что его осуществляет каналообразующий ионофор.
Примером ионофора такого типа является грамицидин А. Он представляет собой линейный полипептид, состоящий из 15 аминокислотных остатков, все они имеют гидрофобные боковые цепи. Две молекулы грамицидина, вероятно, объединяются в бислое и формируют трансмембранный канал, позволяющий моновалентным катионам (Н+ наиболее легко, К+ менее, a Na+ с трудом) перетекать по их электрохимических градиентам. Подобные димеры нестабильны: они постоянно образуются и диссоциируют, так что время, в течение которого канал открыт, составляет в среднем 1 с. При наличии большого электрохимического градиента грамицидин А поможет пропустить около 20 000 катионов в расчете на один открытый канал за 1 миллисекунду, что в 1000 раз больше, чем может перенести за то же время одна молекула подвижного переносчика. Грамицидин - это антибиотик, вырабатываемый определенными штаммами бактерий для уничтожения других микроорганизмов. Его антибактериальное действие основано на том, что он нарушает нормальные градиенты концентраций Н+, Na+ и К+, чрезвычайно важные для жизнедеятельности клеток.
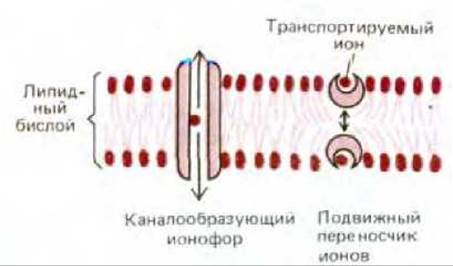
Рис. 6-66. Подвижный переносчик ионов и каналообразующий ионофор. В обоих случаях поток ионов проходит через мембрану только по электрохимическому градиенту.

Рис. 6-67. Молекула валиномицина, связанная с расположенным в центре кольцевой структуры ионом К+ при помощи шести атомов кислорода.
Заключение
Липидные бислои в значительной степени не проницаемы для большинства полярных молекул. Для транспортировки малых водорастворимых молекул в клетку или из клетки в плазматических мембранах содержится большое число различных транспортных белков, каждый из которых ответствен за перенос определенного вещества через мембрану. Существуют два класса мембранных транспортных белков - переносчики и каналы. И те и другие формируют сквозные транспортные пути через липидный бислой.
Белки-переносчики связывают специфические вещества и переносят их через бислой, подвергаясь ряду конформационных изменений, позволяющих экспонировать связывающие вещество участки последовательно: сначала с одной стороны мембраны, а затем с другой. Некоторые белки-переносчики транспортируют вещества только «с горки», другие же, испытывая ряд конформационных изменений, вызываемых гидролизом АТР или связыванием ионов, способны работать как насосы, активно качая связывающееся с ними растворенное вещество «в горку» против его электрохимического градиента.
Белки-каналы образуют в бислое заполненные водой поры, позволяя, таким образом, неорганическим ионам подходящего размера и заряда перемещаться через мембрану по их электрохимическим градиентам. Скорость прохождения в этом случае по крайней мере в 1000 раз выше, чем при транспорте с помощью белков-переносчиков. Эти ионные каналы имеют «ворота» и обычно открываются на короткое время в ответ на специфические возбуждения в мембране, такие, как связывание нейротрансмиттеров (нейротрансмиттер-зависимые воротные каналы) или изменение мембранного потенциала (потенциал-зависимые воротные каналы).