Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Внутриклеточная сортировка макромолекул и сохранение клеточных компартментов
Компартментация в клетках высших организмов
В этом вводном разделе мы дадим краткий обзор клеточных органелл и взаимоотношений между ними. Мы рассмотрим методы, с помощью которых можно проследить движение белков между компартментами, и перечислим основные способы перемещения молекул белка из одного компартмента в другой.
8.1.1. Все эукариотические клетки содержат набор основных ограниченных мембраной органелл [1]
Многие важные биохимические процессы протекают внутри мембран или на их поверхностях. Например, при окислительном фосфорилировании и при фотосинтезе требуется полупроницаемая мембрана для сопряжения транспорта протонов с синтезом АТР. Более того, мембраны служат каркасом для синтеза своих собственных компонентов. Внутренние мембраны эукариотической клетки делают возможной функциональную специализацию различных мембран, что является, как мы увидим, решающим фактором в разделении множества различных процессов, протекающих в клетке.
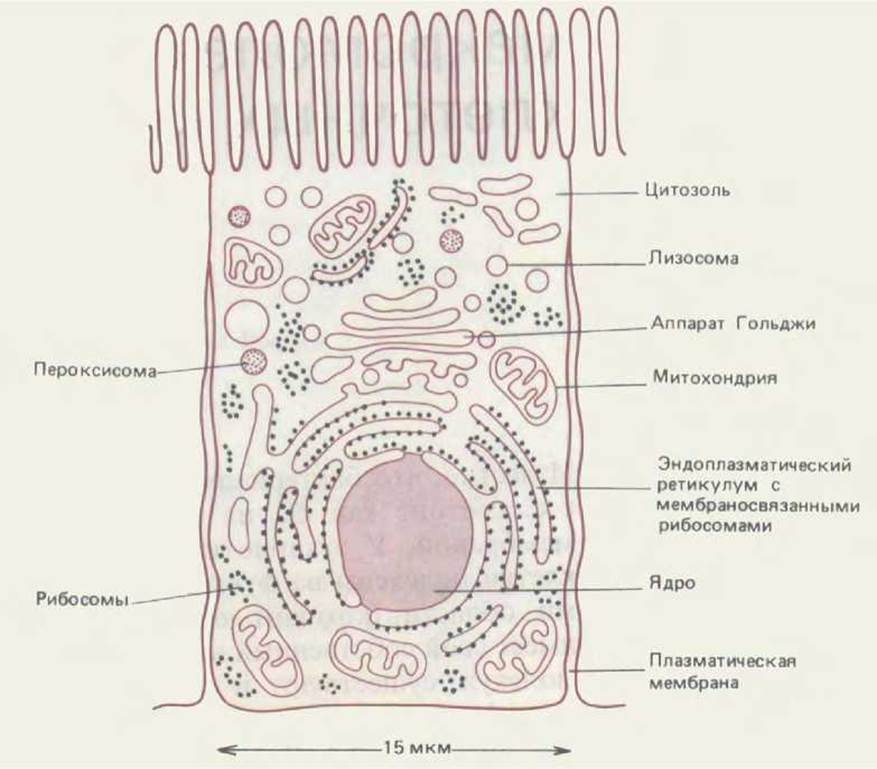
Рис. 8-1. Схематическое изображение основных внутриклеточных компартментов типичной животной клетки. Цитозоль, эндоплазматический ретикулум, аппарат Гольджи, ядро, митохондрия, эндосома, лизосома и пероксисома представляют собой индивидуальные компартменты, отделенные от остальной клетки по крайней мере одной избирательно проницаемой мембраной.
Внутриклеточные компартменты, общие для всех эукариотических клеток, показаны на рис. 8-1. Ядро содержит основную часть генома и является главным местом синтеза ДНК и РНК. Окружающая ядро цитоплазма состоит из цитозоля и расположенных в нем цитоплазматических органелл. Объем цитозоля составляет чуть больше половины от общего объема клетки. Именно в нем синтезируется белок и протекает большинство реакций так называемого промежуточного обмена - т. е. реакций, в которых одни малые молекулы разрушаются, а другие образуются, обеспечивая необходимые строительные блоки для синтеза макромолекул. Около половины всех мембран клетки ограничивают похожие на лабиринт полости эндоплазматического ретикулума (ЭР). На обращенной к цитозолю стороне ЭР находится множество рибосом. Эти рибосомы заняты синтезом интегральных мембранных белков и растворимых белков, предназначенных для секреции или для других органелл. В ЭР также синтезируются липиды для всей остальной клетки. Аппарат Гольджи состоит из правильных стопок уплощенных мембранных мешочков, называемых цистернами Гольджи; он получает из ЭР белки и липиды и отправляет эти молекулы в различные пункты внутри клетки, попутно подвергая их ковалентным модификациям. Митохондрии и хлоропласти растительных клеток производят большую часть АТР, используемого в реакциях биосинтеза, требующих поступления свободной энергии. Лизосомы содержат пищеварительные ферменты, которые разрушают отработанные органеллы, а также частицы и молекулы, поглощенные клеткой извне путем эндоцитоза. На пути к лизосомам поглощенные молекулы и частицы должны пройти серию органелл, называемых эндосомами. Наконец, пероксисомы (известные также как микротельца) представляют собой маленькие пузырьки, содержащие множество окислительных ферментов.
Таблица 8-1. Относительные объемы главных внутриклеточных компартментов в типичной клетке печени (гепатоците)
|
Внутриклеточный компартмент |
Проценты от общего объема клетки1) |
Приблизительное число на одну клетку1) |
|
Цитозоль |
54 |
1 |
|
Митохондрии |
22 |
1700 |
|
Цистерны |
||
|
шероховатого ЭР |
9 |
1 |
|
Цистерны гладкого |
||
|
ЭР плюс цистерны аппарата Гольджи |
6 |
|
|
Ядро |
6 |
1 |
|
Пероксисомы |
1 |
400 |
|
Лизосомы |
1 |
300 |
|
Эндосомы |
1 |
200 |
1) Считается, что все цистерны шероховатого и гладкого ЭР связаны между собой и образуют единый большой компартмент. Напротив, аппарат Гольджи в каждой клетке состоит из множества отдельных наборов собранных в стопки цистерн, и степень взаимосвязи между этими стопками до конца не выяснена.
Кроме только что перечисленных главных мембранных органелл, клетка содержит множество мелких пузырьков, служащих переносчиками веществ между органеллами, а также пузырьков, связывающихся с плазматической мембраной в процессах эндоцитоза и секреции.
В целом, каждая мембранная органелла обладает как свойствами общими для любых клеток, так и специфическими особенностями, связанными со специализированными функциями дифференцированных клеток. Все вместе мембранные органеллы занимают около половины объема клетки (табл. 8-1). Вполне естественно, что для их построения требуется большое количество внутриклеточных мембран. В двух типах клеток млекопитающих, проанализированных для примера в табл. 8-2, общая поверхность мембран эндоплазматического ретикулума превышает поверхность плазматической мембраны соответственно в 25 и в 12 раз. Если говорить о массе и площади, то плазматическая мембрана в большинстве эукариотических клеток - всего лишь небольшая часть всех мембран (рис. 8-2).
Таблица 8-2. Относительные количества мембран разных типов в двух эукариотических клетках
|
Тип мембраны |
Процент от общего количества клеточных мембран |
|
|
Печень Гепатоцит1) |
Поджелудочная железа Экзокринная клетка1) |
|
|
Плазматическая мембрана |
2 |
5 |
|
Мембрана шероховатого ЭР |
35 |
60 |
|
Мембрана гладкого ЭР |
16 |
<1 |
|
Мембрана аппарата Гольджи |
7 |
10 |
|
Митохондрии |
||
|
Внешняя мембрана |
7 |
4 |
|
Внутренняя мембрана |
32 |
17 |
|
Ядро |
||
|
Внутренняя мембрана |
0,2 |
0,7 |
|
Мембрана секреторных пузырьков |
не определен |
о 3 |
|
Мембрана лизосом |
0,4 |
не определено |
|
Мембрана пероксисом |
0,4 |
- » - |
|
Мембрана эндосом |
0,4 |
- » - |
1) Эти клетки сильно различаются по размерам: средний объем гепатоцита около 5000 мкм3, а экзокринной клетки поджелудочной железы около 1000 мкм3. Общая площадь клеточных мембран составляет примерно 110000 мкм2 и 13000 мкм2 соответственно.

Рис. 8-2. Электронная микрофотография участка поперечного среза клетки печени. Отмечены основные внутриклеточные компартменты. (С любезного разрешения D. S. Friend.)
8.1.2. Топология мембранных органелл связана с их эволюционным происхождением [2]
Чтобы понять взаимоотношения между клеточными компартментами. полезно представить себе, как они могли возникнуть в процессе эволюции. Считается, что предшественниками эукариотических клеток были организмы, напоминающие бактерии. У бактерий, как правило, нет внутренних мембран; соответствующие функции у них выполняет плазматическая мембрана. К таким функциям относится, например, транспорт ионов, синтез АТР и синтез липидов. Однако размеры современных эукариотических клеток превышают размер типичной бактериальной клетки (такой, как E.coli) в 10-30 раз. Изобилие внутренних мембран в клетках эукариот можно рассматривать как адаптацию к этому увеличению размеров. По-видимому, плазматической мембраны просто не хватает для поддержания многих жизненно важных функций эукариотической клетки.
Эволюция внутренних мембран, очевидно, шла параллельно со специализацией их функций. У некоторых современных бактерий есть такие участки плазматической мембраны, на которых определенные мембранные белки собраны вместе для выполнения ряда взаимосвязанных функций (рис. 8-3, А). В качестве примера можно привести «пурпурные мембраны» Halobacterium, содержащие бактериородопсин, и хроматофоры фотосинтезирующих бактерий. И те и другие можно назвать примитивными органеллами. У некоторых фотосинтезирующих бактерий эти участки преобразовались в глубокие впячивания плазматической мембраны (рис. 8-3, Б); есть и такие, у которых эти впячивания полностью отшнуровались и превратились в замкнутые мембранные пузырьки, предназначенные для фотосинтеза. Внутренняя поверхность этих пузырьков топологически эквивалентна внешней поверхности клетки (рис. 8-3, В).
Можно предположить, что эукариотическая органелла, возникшая в результате событий, представленных на рис. 8-3, тоже будет иметь внутреннюю поверхность, топологически эквивалентную внешней поверхности клетки. Именно так обстоит дело в случае ЭР, аппарата Гольджи, эндосом и лизосом, а также многих промежуточных пузырьков, участвующих в эндоцитозе и секреции (см. разд. 8.6).
Как уже обсуждалось в гл. 7, митохондрии и хлоропласты отличаются от других окруженных мембраной органелл тем, что имеют свои собственные геномы. Природа этих геномов и близкое сходство белков митохондрий и хлоропластов с белками некоторых современных бактерий подтверждает гипотезу о том, что эти органеллы произошли от бактерий, которые были захвачены другими клетками и первое время существовали в симбиозе с ними (см. разд. 7.5.16). Согласно гипотетической схеме, приведенной на рис. 8-4, А, внутренняя мембрана митохондрий и хлоропластов соответствует исходной плазматической мембране бактерий, а матрикс этих органелл произошел из бактериальной цитоплазмы. Таким образом, эти две органеллы оказались изолированы от путей транспорта, связывающих полости большинства органелл друг с другом и с внеклеточным пространством.
Происхождение клеточного ядра, имеющего особенным образом устроенную двойную мембрану, более загадочно. Известно, что единственная бактериальная хромосома прикреплена к совершенно определенным участкам с внутренней стороны прокариотической плазматической мембраны. Одно из предположений состоит в том, что двуслойная ядерная оболочка могла образоваться из глубокого впячивания плазматической мембраны, как показано на рис. 8-4, Б. Эта гипотеза объясняет, почему внутреннее пространство ядра топологически эквивалентно цитозолю. Действительно, во время митоза у высших эукариот ядерная оболочка разрушается, и содержимое ядра полностью смешивается с цитозолем, чего никогда не происходит ни с одной другой мембранной органеллой. Таким образом, во время митоза клетка временно возвращается к прокариотическому состоянию, когда хромосомы не имеют отдельного компартмента.
Эта эволюционная схема делит основные внутриклеточные компартменты эукариот на пять групп: 1) ядро и цитозоль, связанные между собой ядерными порами и поэтому являющиеся топологически неразрывными (хотя функционально различными); 2) митохондрии; 3) хлоропласты (встречаются только у растений); 4) пероксисомы (их эволюционное происхождение мы обсудим в разд. 8.5) и 5) остальные мембранные органеллы (ЭР, аппарат Гольджи, эндосомы и лизосомы).
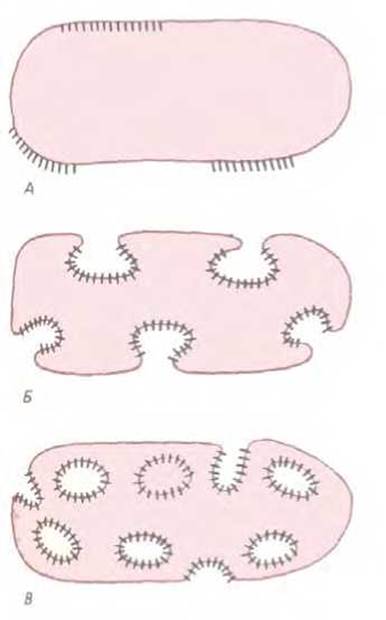
Рис. 8-3. Некоторые специализированные мембраны бактерий. А. Участки поверхности клеточной мембраны, состоящие из скоплений специализированных мембранных белков. Б. Впячивание таких участков увеличивает поверхность мембран, приспособленных для выполнения функций, например, фотосинтеза. В. Внутренняя поверхность образовавшихся в результате впячивания мембранных пузырьков топологически эквивалентна внешней поверхности клетки. Ограниченные мембранами пузырьки существуют у некоторых видов фотосинтезирующих бактерий. Их топологическое отношение к клеточной поверхности подобно отношению ЭР, аппарата Гольджи, эндосом и лизосом к поверхности эукариотических клеток.

Рис. 8-4. Гипотезы эволюционного происхождения митохондрий, хлоропластов, ЭР и клеточного ядра, объясняющие топологические взаимоотношения этих внутриклеточных компартментов в эукариотических клетках. А. Митохондрии и хлоропласты могли возникнуть при поглощении бактерий эукариотической клеткой. С помощью этой гипотезы можно объяснить, почему полость перечисленных выше органелл остается изолированной от обширного везикулярного транспорта, связывающего полости многих других внутриклеточных компартментов. Б.
Возможный путь эволюции ЭР и клеточного ядра. В некоторых бактериях ДНК присоединена к впячиванию плазматической мембраны, называемому мезосомой. Подобное впячивание у очень древней прокариотической клетки могло привести к образованию оболочки вокруг ДНК при сохранении доступа ДНК к цитозолю (так как ДНК должна управлять синтезом белка). Предполагается, что эта оболочка может полностью оторваться от плазматической мембраны, образуя ядерный компартмент, окруженный двойной мембраной. Как показано на рисунке, ядерная оболочка формируется при участии волокнистой структуры — ядерной ламины, и пронизана каналами, называемыми ядерными порами. Благодаря этим порам полость ядра топологически эквивалентна цитозолю. Полость ЭР переходит в пространство между внешней и внутренней ядерными мембранами и топологически эквивалентна внеклеточному пространству.
Внутренние пространства органелл, входящих в последнюю группу, связаны друг с другом и с внеклеточным пространством при помощи транспортных пузырьков, которые отпочковываются от одной органеллы и сливаются с другой. Нужно сказать, что внутренние пространства этих органелл топологически эквивалентны друг другу и внеклеточному пространству и представляют собой функционально связанные части единого комплекса.
8.1.3. Внутриклеточный транспорт белков по ЭР и аппарату Гольджи можно проследить с помощью радиоавтографии [3]
Методы изучения транспорта белков внутри клетки были разработаны в 60-е гг. и впервые применены для наблюдения транспорта из ЭР во внеклеточное пространство в секреторных клетках поджелудочной железы.
Специализированные секреторные клетки, такие, как ацинарные клетки поджелудочной железы, содержат большие количества белкового секрета, заключенного в секреторные везикулы (пузырьки). При стимуляции клетки внешним сигналом содержимое этих пузырьков быстро выбрасывается во внеклеточное пространство путем экзоцитоза. Этот процесс известен как регулируемая секреция. Его следует отличать от конститутивной секреции, представляющей собой другую форму экзоцитоза, происходящего постоянно, в отсутствие стимулирующего сигнала. Ацинарные клетки поджелудочной железы секретируют различные пищеварительные ферменты (амилазу, липазу, дезоксирибонуклеазу и рибонуклеазу), а также предшественники ферментов, так называемые зимогены (например, трипсиноген и химотрипсиноген). Эти предшественники активируются в результате их специфического расщепления протеазами.
Благодаря тому, что группа белков, синтезируемых в ацинарных клетках поджелудочной железы, предназначена для секреции, мы можем судить о пути их передвижения от места синтеза к месту высвобождения. Этот путь можно проследить, сочетая радиоавтографию с электронной микроскопией. Схема соответствующего эксперимента представлена на рис. 8-5. Если клетки кратковременно проинкубировать с [3Н]-аминокислотами (импульсное мечение), а затем различное время выращивать в нерадиоактивной среде, то новосинтезированные белки в первую очередь выявляются в ЭР, а затем в аппарате Гольджи (рис. 8-6). Позднее [3Н]-белки обнаруживаются в больших, незрелых секреторных везикулах вблизи стопки Гольджи (рис. 8-7), где они все более концентрируются; в результате формируются зрелые пузырьки, хорошо различимые на электронных микрофотографиях благодаря их электроноплотному содержимому (рис. 8-7 и 8-8).

Рис. 8-5. Схема, объясняющая, как с помощью электронномикроскопической радиоавтографии можно проследить путь секреторных белков, в состав которых входят [3Н]-меченные аминокислоты. Важно, чтобы все свободные радиоактивные аминокислоты были отмыты до заключения и единственным радиоактивным источником в ткани оставались белки. Затем ткань покрывают тонким слоем фотоэмульсии. Электроны, испускаемые тритием, засвечивают зерна серебра в положениях, несколько смещенных по отношению к источнику излучения, поэтому локализация радиоактивной молекулы определяется менее точно, чем позволяет разрешение электронного микроскопа.
Секреторные везикулы накапливаются в апикальной части ацинарной клетки (т. е. в части, обращенной к системе протоков) между аппаратом
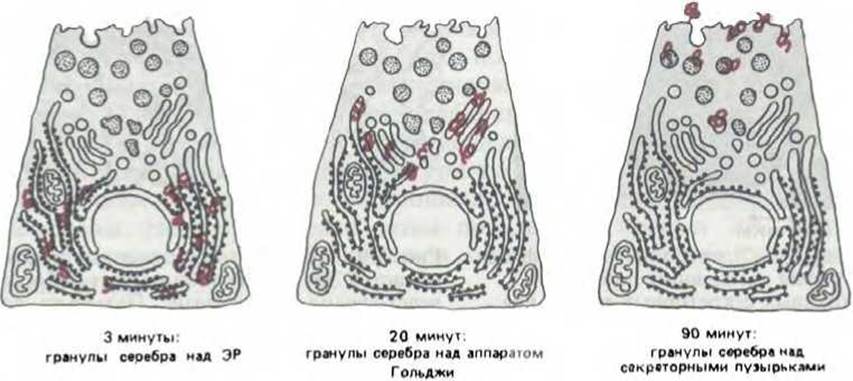
Рис. 8-6. Электронномикроскопическая радиоавтография ацинарных клеток поджелудочной железы, импульсно меченных [3Н]-аминокислотами и затем инкубированных различное время в немеченной среде ("pulse-chase''-метод). С течением времени зерна восстановленного серебра (выделены цветом) перемещаются по направлению к внеклеточному пространству, указывая путь, по которому следуют вновь синтезированные молекулы секреторных белков. Ацинарные клетки необычны тем, что 85% синтезируемых в них белков секретируется; пока клетка не стимулирована к секреции, эти белки сохраняются в секреторных пузырьках.

Рис. 8-7. Электронная микрофотография аппарата Гольджи в ацинарной клетке поджелудочной железы, на которой видны секреторные пузырьки на разных стадиях созревания. Незрелые секреторные пузырьки (называемые в этих клетках конденсирующими вакуолями) отпочковываются от аппарата Гольджи. Секреторные белки в этих пузырьках становятся все более концентрированными (отсюда и название «конденсирующая» вакуоль), что в конечном счете приводит к преобразованию конденсирующих вакуолей в зрелые секреторные пузырьки (вверху слева). Эти секреторные пузырьки в ацинарных клетках поджелудочной железы называют зимогеновыми гранулами. (С любезного разрешения George Palade.)
Гольджи и плазматической мембраной. Когда пузырек сливается с плазматической мембраной, его содержимое попадает во внеклеточное пространство. Эти пузырьки сливаются только с апикальной частью плазматической мембраны, тем самым исключается бесполезное и опасное выделение секрета в межклеточное пространство или в полость других органелл. Более того, экзоцитоз происходит только в ответ на соответствующий внеклеточный химический сигнал, подаваемый нервными клетками или клетками кишечника тогда, когда для пищеварения требуются панкреатические ферменты. Изучено множество секреторных белков из различных типов клеток. Оказалось, что все они проходят один и тот же путь: рибосома → ЭР → аппарат Гольджи → внеклеточное пространство. Те белки, которые должны остаться в плазматической мембране, лизосомах или цистернах аппарата Гольджи, тоже сначала попадают в ЭР и затем оттуда через аппарат Гольджи направляются к местам конечной локализации.
Для выявления более тонких деталей метаболических путей, начинающихся в ЭР, необходимы более сложные методы. Некоторые из попавших в ЭР белков остаются там для выполнения ферментативных функций. Большинство же белков упаковывается в транспортные пузырьки (обычно 50-100 нм в диаметре), которые затем отшнуровываются в специальных местах мембран ЭР, называемых переходными элементами (рис. 89), и специфически сливаются с ближайшей цистерной аппарата Гольджи. После слияния мембрана пузырька становится частью мембраны аппарата Гольджи, а растворимые белки содержимого пузырька доставляются в полость цистерны Гольджи (рис. 8-10). Таким способом растворимые белки избирательно переносятся из одного окруженного мембраной компартмента в другой без проникновения сквозь мембраны. Полагают, что с помощью такого же механизма отделения и слияния пузырьков белки переносятся из одной цистерны Гольджи в другую, и затем к различным местам конечного назначения, в соответствии с их различными функциями. На этом пути белки проходят через целую серию замкнутых пространств, топологически эквивалентных друг другу и внеклеточному пространству (рис. 8-11). На каждом этапе метаболизма белки совершают выбор - остаться в цитозоле или проникнуть в ЭР, остаться в ЭР или поступить в аппарат Гольджи, стать содержимым транспортных пузырьков, направляющихся к лизосомам, или вместо этого попасть в пузырьки, перемещающиеся к клеточной поверхности, и так далее. Выбор в каждом случае зависит от аминокислотной последовательности молекулы белка, содержащей сигналы сортировки, которые определяют путь белка внутри клетки.

Рис. 8-8. Электронная микрофотография очищенного препарата крупных секреторных пузырьков. Такие пузырьки встречаются в специализированных секреторных клетках. (С любезного разрешения Daniel S. Friend.)
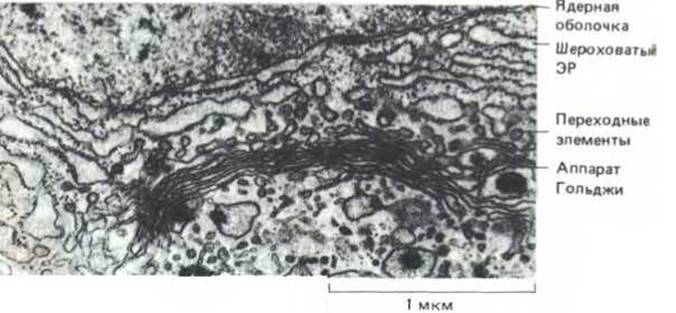
Рис. 8-9. Транспортные пузырьки отпочковываются от особых областей ЭР, почти свободных от рибосом. Эти области называются переходными элементами. Пузырьки сливаются с ближайшей цистерной Гольджи, перенося, главным образом, вновь синтезированные белки и липиды от ЭР к аппарату Гольджи. (С любезного разрешения Brij J. Gupta.)

Рис. 8-10. В процессе везикулярного транспорта сохраняется расположение «сторон» мембран. Обратите внимание, что в мембране компартмента-мишени сохраняется исходная ориентация как белков, так и липидов, а растворимые материалы переносятся из просвета в просвет.
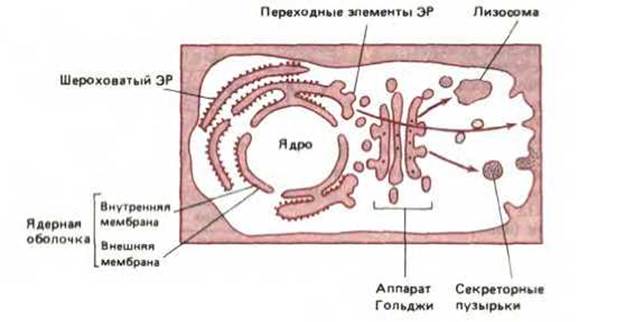
Рис. 8-11. Схема клетки, в которой топологически эквивалентные области выделены цветом. В принципе, циклическое отпочковывание и слияние пузырьков позволяет связать просвет (полость) любой органеллы с другим просветом и с внеклеточным пространством. Однако некоторые органеллы (например, митохондрии и хлоропласты) не связаны с другими органеллами везикулярным транспортом. Таким образом, они изолированы от изображенных здесь транспортных путей между органеллами.
8-3
8.1.4. Транспорт белков происходит по двум основным путям - через цитозоль и через ЭР [4]
Основные пути транспорта белков показаны на рис. 8-12. Практически все белки образуются на рибосомах, расположенных в цитозоле (кроме нескольких, синтезирующихся на митохондриальных рибосомах хлоропластов). Затем их пути расходятся. Белки, принадлежащие одной транспортной ветви, после завершения их синтеза выделяются в цитозоль. Некоторые из них содержат сигналы сортировки, направляющие их из цитозоля в митохондрии, хлоропласты (у растений), ядро или пероксисомы, другие же - их большинство - не имеют специфических сигналов сортировки и остаются в цитозоле в качестве постоянных компонентов.
Другой основной путь транспорта используется при синтезе белков, предназначенных для выведения из клетки, а также белков, которые должны стать компонентами ЭР, аппарата Гольджи, плазматической мембраны или лизосом. Все эти белки по мере их образования переносятся в ЭР при помощи сигналов сортировки, расположенных обычно на N-конце. Рибосомы, на которых собираются такие белки, остаются связанными с мембраной ЭР в течение недолгого времени после начала синтеза полипептидной цепи. Как только очередной участок полипептидной цепи синтезируется, он проникает через липидный бислой этой мембраны. Некоторые белки затем попадают в просвет ЭР, другие остаются частично заключенными в мембрану в качестве трансмембранных белков.
Обычно проходит не больше одной или двух минут с момента высвобождения белка в цитозоль до поступления его в соответствующую органеллу. Белки, предназначенные для ядра, митохондрий или пероксисом, достигнув этих органелл, на этом заканчивают свой путь, между тем белки, попадающие в ЭР, вовлекаются в дальнейший транспорт, который, как полагают, происходит с помощью транспортных пузырьков, отделяющихся от одной мембраны и сливающихся с другой. По-видимому, для достижения конечного пункта должно произойти несколько таких циклов, и путь от мембраны ЭР до места назначения может занять один час.

Рис. 8-12. Упрощенная схема путей метаболизма белков. Сигналы, направляющие движение конкретного белка по определенной «дороге» и, следовательно, определяющие локализацию этого белка в клетке, содержатся в его аминокислотной последовательности. «Путешествие» начинается с синтеза белка на рибосоме и заканчивается его прибытием к месту конечного назначения. На каждой промежуточной стадии (изображены прямоугольниками) принимается решение - оставить данный белок на ней или транспортировать дальше. В принципе, сигнал может требоваться либо для удержания белка в каждом из указанных компартментов, либо для его продвижения дальше, а альтернативное событие может происходить «по умолчанию» (т. е. без специального сигнала). На этой схеме те пути, которые, вероятнее всего, требуют специальных сигналов, выделены цветом; те же пути, которые, скорее всего, выбираются по умолчанию, показаны черными стрелками. Так как образование эндосом изучено плохо, здесь не изображено никаких путей, ведущих к эндосомам.
8.1.5. Белки могут перемешаться между компартментами двумя принципиально различными способами [5]
Чтобы понять общие принципы работы сигналов сортировки, важно различать два совершенно различных пути, по которым белки перемещаются из одного компартмента в другой. Во-первых, они могут непосредственно проникать через мембрану, попадая из пространства, топологически эквивалентного цитозолю, в пространство, топологически эквивалентное внеклеточному, или наоборот. Этот путь требует наличия в мембране специального белка-транслокатора, кроме того, молекула транспортируемого белка должна развернуться, чтобы, подобно змее, «проползти» сквозь мембрану. В качестве примера такого рода событий может служить перемещение определенных белков из цитозоля в просвет ЭР. Второй путь передвижения белковых молекул опосредован транспортными пузырьками. Эти пузырьки захватывают определенные молекулы в полости одного компартмента (от которого они отшнуровываются) и переносят их в другой компартмент, сливаясь с ним. Именно так происходит перенос растворимых белков из ЭР к аппарату Гольджи (см. рис. 8-10). При таком везикулярном транспорте белки не пересекают никаких мембран, поэтому они переносятся только между компартментами, топологически эквивалентными друг другу.
Оба типа транспортных процессов избирательно контролируются с помощью специальных белков, выполняющих роль сигналов сортировки. У белка, который напрямую переносится через мембрану, эти сигналы распознаются транслокатором в мембране. А в транспортный пузырек белок попадает, если его сигнал сортировки связывается с рецептором на мембране пузырька. Вероятно, существуют и такие транспортные пузырьки, которые способны захватывать белки, утерявшие специфические сигналы сортировки. В любом случае вновь образующиеся пузырьки переносят только предназначенные для этого белки.
В настоящее время некоторые сигналы сортировки в составе белков известны, тогда как большинство комплементарных им мембранных рецепторов - нет. Кроме того, мы не знаем почти ничего о транспортных пузырьках. Можно предположить только, что их существует множество видов, в соответствии с различными переносами, которые они должны осуществлять. Чтобы выполнять свои функции, каждый пузырек должен захватывать только определенные белки и сливаться только с определенной мембраной-мишенью: например, пузырек, переносящий молекулы от ЭР к аппарату Гольджи, не должен включать белки, которые останутся в ЭР, и должен сливаться только с аппаратом Гольджи, а не с другими органеллами. В этой главе мы рассмотрим, каким образом транспортные пузырьки достигают такой избирательности.
8.1.6. Сигнальные пептиды и сигнальные участки определяют судьбу белка [6]
Полагают, что на белках существуют два типа сигналов сортировки, направляющие их, шаг за шагом, вдоль разветвляющихся путей (рис. 8-12). Для некоторых стадий сигналы сортировки представляют собой протяженный участок аминокислотной последовательности длиной 15-60 остатков. Когда эта стадия пройдена, такой сигнальный пептид отрезается. Сигналом сортировки для других стадий, вероятно, служит определенная трехмерная структура, образуемая атомами поверхности белка при свертывании его молекулы. Аминокислотные остатки, формирующие такие сигнальные участки, могут быть расположены очень далеко друг от друга в линейной последовательности белка (рис. 8-13). Сигнальные пептиды направляют белки из цитозоля в ЭР, митохондрии, хлоропласты и ядро; они также отвечают за то, чтобы некоторые белки остались в ЭР. Сигнальные участки, видимо, играют важную роль при распознавании определенных лизосомных белков специальным ферментом в аппарате Гольджи.
Чтобы выяснить пункт назначения того или иного белка внутри клетки, необходимо определить тип его сигнального пептида (табл. 8-3). Белки, которые должны попасть в ЭР, обычно несут N-концевой сигнальный пептид. Его центральная часть образована 5-10 гидрофобными аминокислотными остатками. Большинство этих белков направляется из ЭР в аппарат Гольджи; те же, которые имеют на С-конце специфическую последовательность из четырех аминокислот, остаются в качестве постоянных компонентов. Многие белки, предназначенные для митохондрий, имеют сигнальные пептиды, в которых положительно заряженные аминокислотные остатки чередуются с гидрофобными. Среди белков, направляющихся в ядро, большинство имеет сигнальные пептиды, образованные кластером положительно заряженных аминокислотных остатков. Наконец, некоторым белкам цитозоля присущи сигнальные пептиды, с которыми ковалентно связывается жирная кис-

Рис. 8-13. Два способа образования на белке транспортного сигнала. А. Сигнал представляет собой единственный дискретный отрезок аминокислотной последовательности, называемый сигнальным пептидом. В свернутом белке сигнальный пептид располагается снаружи. Сигнальные пептиды чаще находятся на конце полипептидной цепи (как показано здесь), но могут быть и в других местах. Сигнальные пептиды выявляют экспериментально по их эффекту на внутриклеточную сортировку белков. Для этого сигнальный пептид с помощью методов рекомбинантных ДНК «пришивают» к какому-нибудь другому белку. Б. Сигнальный участок может быть сформирован при наложении аминокислот из удаленных друг от друга районов при сворачивании молекулы; или же такой сигнал образуется из отдельных «пятен» на поверхности свернутого белка, удаленных друг от друга на известное расстояние. И в том, и в другом случае транспортный сигнал зависит от трехмерной конформации белка. По этой причине точно локализовать такие сигналы чрезвычайно трудно.
Таблица 8-3. Типичные последовательности сигнальных пептидов

Заряженные остатки обозначены + или —. Большой блок гидрофобных остатков заключен в рамку. H3N-обозначает N-конец белка; — COO' обозначает карбоксильный конец лота, направляющая эти белки к мембранам без проникновения в ЭР.
Роль каждого такого сигнального пептида в доставке белка по месту назначения была продемонстрирована в опытах по генной инженерии. Например, помещение N-концевого сигнального пептида, связывающегося с ЭР, в начало белка цитозоля, заставляло этот белок направляться к ЭР. Сигнальные пептиды белков, имеющих одно и то же конечное назначение, функционально взаимозаменяемы, хотя их аминокислотные последовательности могут сильно различаться. Возможно, для процесса распознавания сигнала физические свойства (такие, например, как гидрофобность) оказываются важнее, чем точная последовательность аминокислот.
Сигнальные участки анализировать гораздо труднее, чем сигнальные пептиды, поэтому об их структуре известно гораздо меньше. Поскольку они образуются из сложной трехмерной конфигурации свернутого белка, их невозможно просто перенести от одного белка к другому. Более того, экспериментальное повреждение сигнального участка часто влечет за собой повреждение белка как целого.
8.1.7. Клетки не могут строить свои мембранные органеллы de novo: им требуется информация, содержащаяся в самой органелле [7]
Когда клетка воспроизводится и делится, она должна удваивать свои мембранные органеллы. Обычно это происходит путем увеличения размеров этих органелл при включении в них новых молекул; увеличенные органеллы затем делятся и распределяются по двум дочерним клеткам. Вряд ли клетка в состоянии создавать эти органеллы de novo. Если, например, из нее полностью удалить ЭР, она не сумеет его реконструировать. Ведь мембранные белки, определяющие специфику ЭР и выполняющие множество его ключевых функций, сами являются продуктами ЭР: без ЭР или по крайней мере без мембраны, содержащей транслокаторы, необходимые для импорта белков в ЭР (и при отсутствии транслокаторов, которые требуются для импорта белков в другие органеллы), новый ЭР не может быть построен.
Следовательно, для формирования мембранных органелл недостаточно только информации ДНК, определяющей белки органелл. Необходима также «эпигенетическая» информация в виде хотя бы одного характерного белка в мембране органеллы. Эта информация передается от родительской клетки потомству с самой органеллой. Вероятно, такая информация необходима для поддержания компартментации клетки, тогда как информация, содержащаяся в ДНК, необходима для «размножения» нуклеотидных и аминокислотных последовательностей.
Заключение
Эукариотические клетки содержат внутриклеточные мембраны, замыкающие около половины общего объема клетки в отдельные внутриклеточные компартменты. Основные виды мембранных органелл во всех эукариотических клетках следующие: эндоплазматический ретикулум, аппарат Гольджи, ядро, митохондрии, лизосомы, эндосомы и пероксисомы; растительные клетки содержат также хлоропласты. Каждая органелла имеет в своем составе различные белки, определяющие ее уникальные функции.
Каждый вновь синтезированный белок органелл проходит от рибосомы до органеллы особый путь, определяемый либо сигнальным пептидом, либо сигнальным участком. Сортировка белков начинается с первичной сегрегации, при которой белок либо остается в цитозоле, либо переносится в другой компартмент (например, в ядро, митохондрию или эндоплазматический ретикулум). Белки, попадающие в ЭР, претерпевают дальнейшую сортировку по мере того, как они переносятся в аппарат Гольджи и затем из аппарата Голъджи в лизосомы, в секреторные пузырьки или к плазматической мембране. Некоторые белки остаются в ЭР и в различных цистернах аппарата Гольджи. Белки, предназначенные для других компартментов, видимо, попадают в транспортные пузырьки, которые отшнуровываются от одного компартмента и сливаются с другим.