Молекулярная биология клетки - Том 2 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1993
Внутриклеточная сортировка макромолекул и сохранение клеточных компартментов
Транспорт белков и РНК в ядро и из ядра
Содержимое клеточного ядра (нуклеоплазма) отделено от цитоплазмы ядерной оболочкой. Ядерная оболочка образована двойной мембраной. Сферическая внутренняя ядерная мембрана содержит специфические белки, выступающие в качестве сайтов связывания ядерной ламины, которая поддерживает мембрану и контактирует с хромосомами и ядерными РНК. Эта мембрана окружена внешней ядерной мембраной, очень схожей с мембраной эндоплазматического ретикулума, в которую она переходит (рис. 8-19). Внешнюю (наружную) ядерную мембрану можно рассматривать как особую часть мембраны ЭР. Подобно мембранам шероховатого ЭР (см. разд. 8.6.1), внешняя ядерная мембрана усеяна рибосомами, участвующими в синтезе белка. Белки, образованные на этих рибосомах, переносятся в пространство между внешней и внутренней ядерными мембранами (перинуклеарное пространство), которое в свою очередь связано с просветом ЭР (см. рис. 8-19).
Ядро содержит множество белков, необходимых для обеспечения его уникальных функций. Эти белки (к ним относятся гистоны, ДНК-и РНК-полимеразы, белки-регуляторы различных генов и белки, участвующие в процессинге РНК) синтезируются в цитозоле и затем попадают в ядро. Чтобы достигнуть внутреннего пространства ядра (просвета ядра), они должны пройти внешнюю и внутреннюю ядерные мембраны. Этот транспорт происходит избирательно: многие белки, образованные в цитозоле, никогда не попадают в ядро.
8.3.1. Двойную ядерную мембрану пронизывают ядерные поры [16]
У всех эукариот, от дрожжей до человека, ядерная оболочка пронизана ядерными порами. Поры окружены большими кольцевыми структурами, называемыми поровыми комплексами (их внутренний диаметр составляет ол. М ~ 80 нм, а масса - 50-100 млн. Каждый комплекс образован набором больших белковых гранул, сгруппированных в октагональную структуру (рис. 8-20, А и 8-21). Поровый комплекс пронизывает двойную мембрану, связывая по окружности поры липидный бислой внутренней и внешней мембран в единое целое (рис. 8-20, Б). Несмотря на эту непрерывность, которая должна была бы обеспечивать диффузию компонентов между внешней и внутренней мембранами, они остаются химически различными.
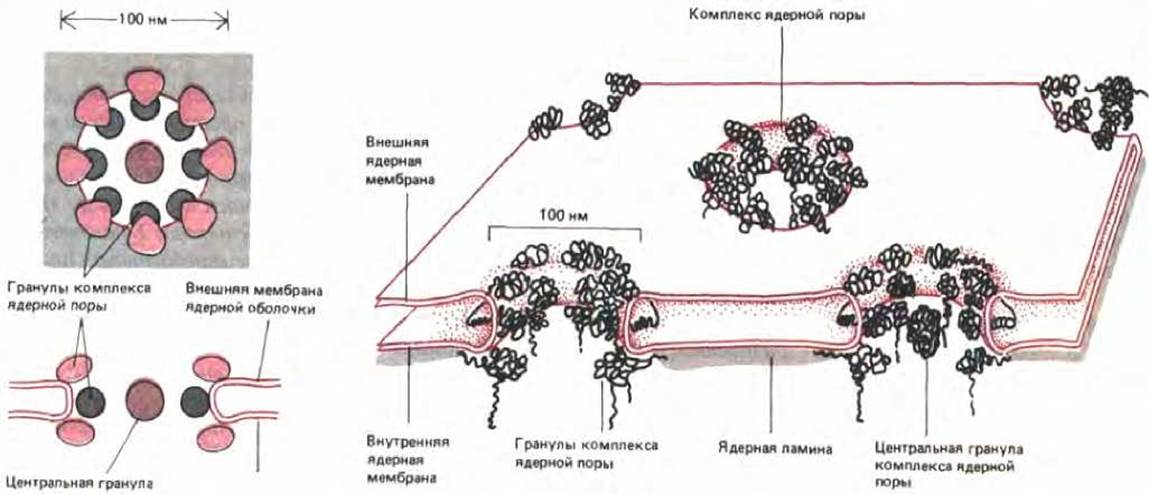
Рис. 8-20. Схема, показывающая расположение ядерных поровых комплексов в ядерной оболочке Вид сверху и поперечное сечение по центру поры (слева). Выделенная цветом «центральная гранула» присутствует в одних порах и отсутствует в других; эти гранулы могут являться частью поры, но не исключено, что это просто крупные частицы, зафиксированные в момент прохождения перового канала. Схема небольшого участка ядерной оболочки (справа).

Рис. 8-21. Электронная микрофотография препарата ядерных поровых комплексов (негативное контрастирование). Каждую пору окружает кольцо из восьми гранул. Размер каждой гранулы примерно равен размеру рибосомы. (С любезного разрешения A. Faberge.)
«Дыра» в центре каждого комплекса (ядерная пора) представляет собой водный канал, сквозь который водорастворимые молекулы курсируют между ядром и цитоплазмой. Часто создается впечатление, что это отверстие «закупорено» большой гранулой, которая, как полагают, состоит из вновь синтезированных рибосом или других частиц, видимых в момент переноса в цитоплазму (см. рис. 8-20, Б). Эффективный размер поры в состоянии покоя был определен с помощью эксперимента, в ходе которого в цитозоль вводили различные меченые молекулы неядерного происхождения и измеряли скорости их диффузии в ядро. Оказалось, что малые молекулы (5 кДа и меньше) проникают в ядро с такой скоростью, что ядерную оболочку можно считать для них свободно проницаемой. Концентрация белка с мол. массой 17 кДа выравнивается между цитоплазмой и ядром за 2 мин; для белка с мол. массой 44 кДа это происходит за 30 мин, а глобулярные белки, имеющие свыше 60 кДа, едва ли вообще проникают в ядро. Количественный анализ подобных данных подтверждает, что ядерный поровой комплекс содержит заполненный водой цилиндрический канал диаметром около 9 нм и длиной 15 нм (рис. 8-22). Эти размеры сравнимы с размером беспорядочно расположенных каналов, которые видны на некоторых электронных микрофотографиях.
По-видимому, ядерная оболочка приспособлена к тому, чтобы закрывать содержимое ядерного компартмента (нуклеоплазму) от множества частиц, филаментов и больших молекул, работающих в цитоплазме. Зрелые цитоплазматические рибосомы, например, слишком велики, чтобы проникать через эти 9-нанометровые каналы, поэтому весь белковый синтез ограничивается цитоплазмой. Непонятно только, как же попадают в ядро большие молекулы, которые там необходимы, например, ДНК- и РНК-полимеразы, имеющие мол. массу субъединиц от 100 до 200 кДа? Недавно получены доказательства того, что эти и многие другие ядерные белки взаимодействуют с белками-рецепторами, расположенными на границе ядерных пор, и эти рецепторы активно переносят большие белки в ядро, увеличивая канал поры.
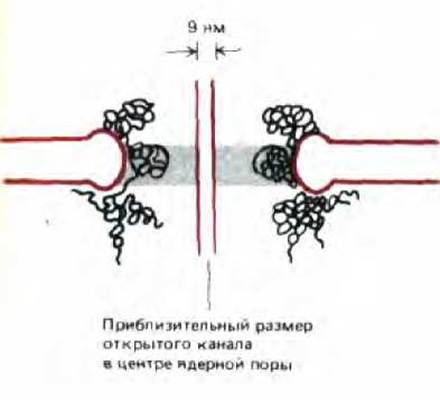
Рис. 8-22. Поперечный разрез ядерной поры (упрощенная схема). Воображаемый цилиндр, изображенный в центре поры, показывает эффективный размер открытого канала, рассчитанный путем измерений транспортируемых частиц. На некоторых электронных микрофотографиях можно различить нити, заполняющие почти все внутреннее пространство поры. Возможно, присутствие этих нитей приводит к уменьшению просвета поры до 9 нм.
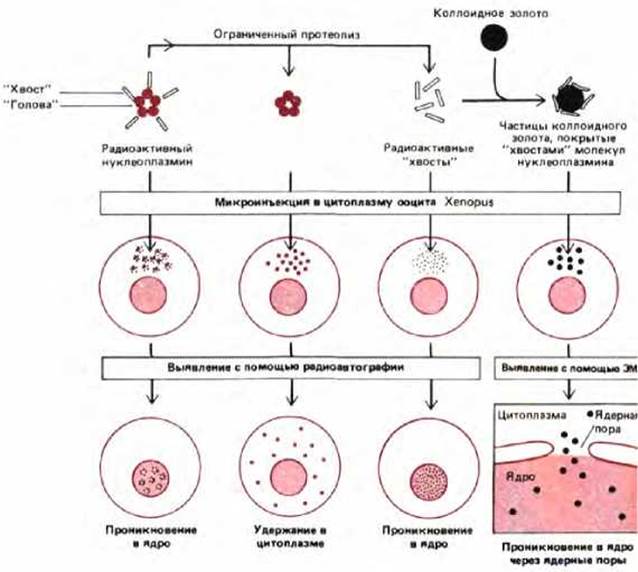
Рис. 8-23. Эксперименты, демонстрирующие проникновение избранных белков в ядро через ядерные поры. Нуклеоплазмин представляет собой большой пентамерный белок с отличающимися «головными» и «хвостовыми» доменами. «Головки» можно отрезать от «хвостов» при помощи ограниченного протеолиза. Интактный нуклеоплазмин при введении его в цитоплазму ооцита шпорцевой лягушки быстро накапливается в ядре, несмотря на то, что он слишком велик, чтобы пассивно проникать сквозь маленький канал в центре перового комплекса. По-видимому, сигнал, направляющий импорт этого белка в ядро, расположен в «хвостовом» домене, потому что введенные в цитоплазму ооцита отдельные «хвосты» переносятся в ядро, а отдельные «головы»-нет. Роль ядерных пор в этом управляемом сигналом переносе показана с помощью электронной микроскопии. Нуклеоплазминовые «хвосты» связывали с частицами коллоидного золота, хорошо видимыми в электронном микроскопе из-за их высокой электроноплотности. Присоединенные нуклеоплазминовые «хвосты» вызывают проникновение частиц коллоидного золота через ядерные поры.
8.3.2. Белки активно проникают в ядро через ядерные поры [17]
Если из ядра экстрагировать белки, а затем с помощью микроинъекции ввести их в цитоплазму, то даже очень крупные белки вновь будут накапливаться в ядре. Одним из наиболее хорошо изученных примеров является мажорный ядерный белок нуклеоплазмин, который можно протеолитически расщепить на «голову» и «хвост». В экспериментах с микроинъекциями «хвостовая» часть проникает в ядро, а «головная»-нет (рис, 8-23). Если «хвосты» связать с частицами коллоидного золота диаметром 20 нм (что гораздо больше внутреннего диаметра находящейся в покое ядерной поры), то частицы золота накапливаются в ядре, их можно видеть в ядерных порах и в процессе транспорта (рис. 8-23 и 8-24). Следовательно, ядерная пора может «открываться» и пропускать такой большой и чужеродный объект, как частица золота. Похоже, что пора работает подобно клапану, который открывается в ответ на сигнал от достаточно крупного белка. Как все это происходит на молекулярном уровне - остается загадкой.
Белки, подобные нуклеоплазмину, активно транспортируются через поры, возможно даже оставаясь при этом в свернутом состоянии. Эксперименты по воссозданию активного ядерного транспорта in vitro подтверждают, что необходимую для этого процесса энергию клетка получает в результате гидролиза АТР.
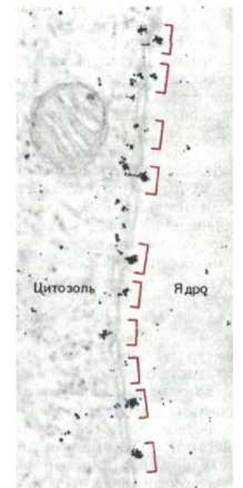
Рис. 8-24. Электронная микрофотография, показывающая прохождение частиц коллоидного золота, покрытых нуклеоплазмином (см. рис. 8-23) в ядро через ядерные поры, локализация которых отмечена цветными скобками. Сходный результат получается, если частицы золота связаны только с хвостовыми частями молекул нуклеоплазмина. Эти частицы больше в диаметре, чем покоящаяся пора. Следовательно, пора должна «открываться», чтобы обеспечить их прохождение в ядро. (По С. Feldherr, E. Kallenbach, N. Schultz, J. Cell Biol. 99: 2216-2222, 1984.)
8-11
8-12
8-13
8.3.3. В ядро активно переносятся только белки, содержащие специальные сигналы [18]
Избирательность ядерного транспорта обеспечивается сигналами ядерного импорта, которые имеются только у ядерных белков. Мы уже говорили о том, что у нуклеоплазмина сигнал такого типа расположен в хвостовой части молекулы. Для некоторых других ядерных белков сигналы ядерного импорта были локализованы более точно с помощью методов генной инженерии. Оказалось, что они могут находиться в любой части молекулы белка, состоят из короткого пептида (обычно от четырех до восьми аминокислотных остатков), обогащенного положительно заряженными аминокислотами лизином и аргинином и обычно содержат пролин. Сигнал такого типа впервые был идентифицирован в белке вируса SV40, так называемом Т-антигене, большом (90 кДа) белке, необходимом для репликации вирусной ДНК в ядре. В норме Т-антиген накапливается в ядре вскоре после его синтеза в цитозоле. Однако замена одной-единственной аминокислоты делает невозможной транспортировку белка и вызывает его накопление в цитоплазме (рис. 8-25). Было высказано предположение, что мутация затрагивает последовательность сигнала ядерного импорта. В ходе дальнейших экспериментов ДНК нормального Т-антигена, кодирующую этот район, состыковали с геном, кодирующим мутантный цитоплазматический белок. При этом была определена минимальная последовательность, обусловливающая способность «гибридного» белка проникать в ядро. Удалось показать, что сигнал ядерного импорта для Т-антигена представляет собой цепочку из восьми следующих друг за другом аминокислот, расположенных во внутреннем районе его полипептидной цепи (см. табл. 8-3). Дальнейшие эксперименты убедили в том, что эта сигнальная последовательность успешно функционирует и в том случае, если ее синтезировать в виде короткого пептида и химически присоединить к любой случайно выбранной боковой цепи лизина в «цитоплазматическом» мутантном белке. Таким образом, локализация сигнала ядерного импорта в белке, по-видимому, не важна.
Механизм транспорта белков в ядро принципиально отличается от механизмов транспорта белков в другие органеллы (которые будут описаны далее). Отличие состоит в том, что транспорт белков в ядро происходит через регулируемые водные поры, а не через одну или более мембран. Более того, когда ядро «разбирается» в митозе, его содержимое перемешивается с цитозолем, и ядерные белки выбрасываются наружу. Когда ядро собирается вновь, группы хромосом сначала упаковываются в собственные двойные мембраны, прилежащие к ним так плотно, что растворимые белки, включая множество прежних компонентов ядра, там «не помещаются». Затем эти заключенные в оболочку хромосомы объединяются, формируя одно ядро, в которое надо транспортировать необходимые белки из цитозоля. Возможно, именно потому, что молекулам ядерного белка предстоит повторный перенос в ядро, сигнальный пептид ядерного импорта не отрезается от них после попадания в ядро. Напротив, когда молекула белка попадает в любую другую мембранную органеллу, она передается от поколения к поколению уже внутри данного компартмента, и ее никогда не приходится транспортировать в него вновь. Поэтому сигнальные пептиды таких молекул удаляются, как только белок переносится внутрь компартмента.
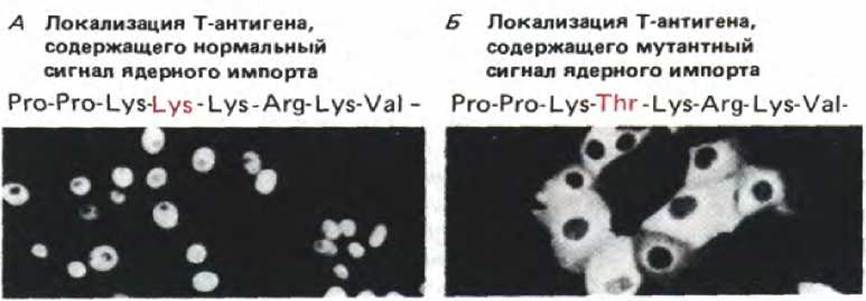
Рис. 8-25. Местонахождение Т-антигена вируса SV40, содержащего или не содержащего сигнальный пептид, определяющий ядерную локализацию. Т-белок дикого типа содержит приведенную здесь богатую лизином последовательность и импортируется в ядро к месту конечного назначения, что показано с помощью иммунофлуоресцентного окрашивания антителами к Т-антигену (А). Т-антигены с измененным сигнальным пептидом (например, с заменой лизина на треонин) остаются в цитоплазме (Б). (По D. Calderon, В. Roberts, W. Richardson, A. Smith, Cell 39: 499-509, 1984.)
8.3.4. Некоторые РНК покидают ядро через ядерные поры [19]
Ядерная оболочка типичной клетки млекопитающих содержит от 3000 до 4000 пор (примерно 11 пор на 1 мкм2 площади мембраны). Если в клетке синтезируется ДНК, то для упаковки в хроматин вновь синтезированных молекул необходимо каждые 3 минуты переносить из цитоплазмы в ядро около 106 молекул гистонов. Это значит, что каждая пора должна пропускать приблизительно 100 молекул гистонов в минуту. Если клетка быстро растет, то каждая пора должна еще пропускать около трех вновь образованных рибосом из ядра в цитоплазму, поскольку рибосомы формируются в ядре, а функционируют в цитозоле (см. разд. 9.4.17). И это лишь очень небольшая часть всего транспорта, проходящего через ядерные поры.
Особый интерес вызывает механизм экспорта новых субъединиц рибосом. Эти частицы слишком велики (около 15 нм в диаметре), чтобы проникать через 9-нанометровые каналы. Более вероятно, что они проникают сквозь ядерные поры, используя систему активного транспорта. Полагают, что и молекулы информационной РНК в составе рибонуклеопротеиновых частиц (в комплексе со специальными белками) переносятся из ядра в цитоплазму активно. Если частицы коллоидного золота диаметром 20 нм, подобные тем, что были использованы в экспериментах с нуклеоплазмином (см. рис. 8-24), связать с молекулами малых РНК (тРНК или 5S-PHK) и затем инъецировать в ядро ооцита лягушки, то они быстро переносятся через ядерные поры в цитоплазму. С другой стороны, если их ввести в цитоплазму ооцита, они останутся там. Видимо, помимо рецепторов, узнающих сигналы ядерного импорта, поры содержат один или более рецепторов, распознающих молекулы РНК (или связанные с ними белки), предназначенные для цитозоля; когда эти рецепторы связаны, пора катализирует активный транспорт наружу вместо транспорта внутрь ядра. Заметим, что хотя некоторые белки ядерных пор (включая мажорный мембранный белок с мол. массой 190 кДа) недавно были выделены, до сих пор неизвестно, как именно работает ядерная пора.
Заключение
Ядро заключено в оболочку, состоящую из двух концентрических мембран. Внешняя ядерная мембрана переходит я мембрану ЭР, а пространство между внешней и внутренней ядерными мембранами переходит в полость ЭР. Молекулы РНК и рибосомы образуются в ядре и переносятся в цитозолъ, тогда как все белки, функционирующие в ядре, синтезируются в цитозоле и переносятся в ядро. Обмен материалом между ядром и цитоплазмой происходит через ядерные поры, обеспечивающие прямой путь сквозь внутреннюю и внешнюю ядерные мембраны.
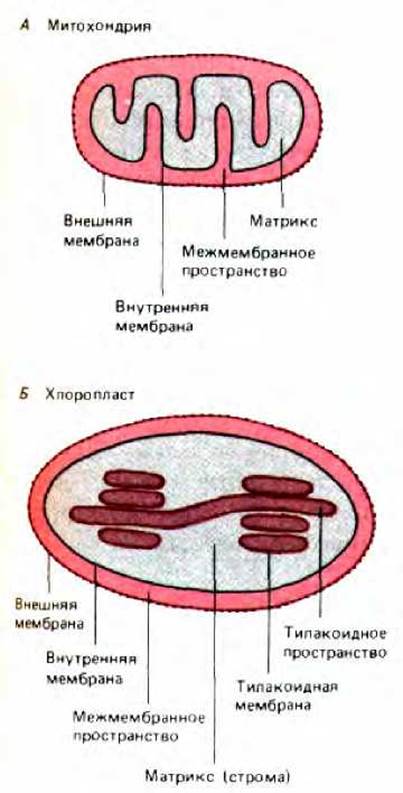
Рис. 8-26. Основные субкомпартменты митохондрий и хлоропластов. Топология хлоропласта может быть получена из топологии митохондрий простым способом: если впячивания внутренней митохондриальной мембраны полностью отпочкуются, то получится компартмент, топологически эквивалентный тилакоидам хлоропластов.
Белки, содержащие сигналы ядерного импорта, активно переносятся в ядро через поры, и узнаются эти белки по коротким, положительно заряженным сигнальным пептидам; поскольку сигнальный пептид не удаляется после переноса, ядерные белки могут быть перенесены в ядро тогда, когда это требуется при сборке ядра после митоза. Молекулы РНК и, возможно, субъединицы рибосом, активно переносятся через поры из ядра в цитоплазму.