Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Иммунная система
Клеточная основа иммунитета
18.1.1. Иммунная система человека состоит из триллионов лимфоцитов [1]
За специфичность иммунитета ответственны лимфоциты - одна из групп лейкоцитов. Они содержатся в больших количествах в крови, в лимфе (бесцветная жидкость лимфатических сосудов, соединяющих между собой лимфатические узлы) и в специализированных лимфоидных органах, таких как тимус (вилочковая железа), лимфатические узлы, селезенка и аппендикс (рис. 18-1).
Общее число лимфоцитов в организме человека составляет около 2-1012; по клеточной массе иммунная система сравнима с печенью или мозгом. Хотя лимфоциты уже давно признаны одним из важных клеточных компонентов крови, их центральная роль в иммунитете была продемонстрирована лишь в конце 50-х годов. В решающих экспериментах мышей или крыс подвергали сильному облучению, приводившему к гибели большинства лейкоцитов, в том числе лимфоцитов. Облученным животным, неспособным к иммунному ответу, можно было вводить клетки различных типов, чтобы выяснить, какие из них восстанавливают иммунную реактивность. Таким свойством обладали только лимфоциты (рис. 182). Поскольку восстанавливались как клеточные формы иммунного ответа, так и выработка антител, полученные результаты доказывали, что лимфоциты ответственны за оба класса иммунных ответов. Когда проводились эти эксперименты, лимфоциты были одним из наименее изученных типов клеток позвоночных; сейчас они относятся к наиболее изученным.

Рис. 18.1. Лимфоидные органы человека. Лимфоциты развиваются в тимусе и костном мозге (на схеме темноокрашенные участки), которые поэтому называют первичными лимфоидными органами. Новообразованные лимфоциты мигрируют из этих первичных органов во вторичные лимфоидные органы (светлоокрашенные участки), где могут реагировать с антигеном. Показаны только некоторые из вторичных лимфоидных органов.
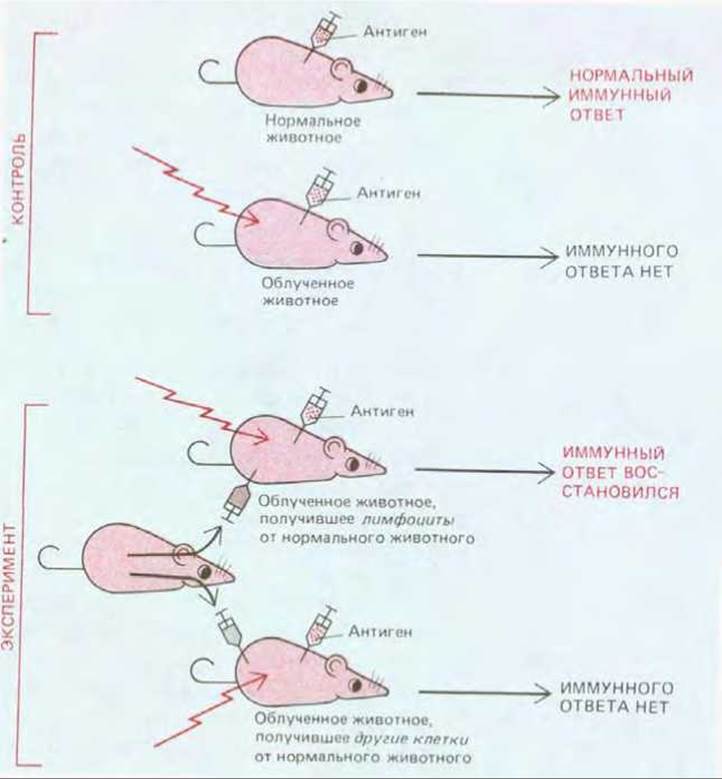
Рис. 18-2. Классический эксперимент, показывающий, что за узнавание чужеродных антигенов и реакцию на них ответственны лимфоциты. Важная особенность всех таких эспериментов с переносом клеток состоит в том, что донор и реципиент принадлежат к одной инбредной линии и поэтому генетически идентичны. Если лимфоциты донора ввести генетически отличающемуся от него животному, которое было подвергнуто облучению, то они будут реагировать против «чужеродных» для них антигенов реципиента и могут вызвать его гибель.
18.1.2. В-лимфоциты реализуют гуморальные иммунные ответы, а Т-лимфоциты - иммунные ответы клеточного типа [2]
В 60-х годах было установлено, что два основных класса иммунных реакций опосредуются двумя различными классами лимфоцитов: Т- клетки, развивающиеся в тимусе, ответственны за клеточный иммунитет, а В-клетки, которые у млекопитающих развиваются в костном мозге взрослой особи или в печени плода, вырабатывают антитела. Такого рода дихотомию лимфоидной системы первоначально удалось выявить у животных с экспериментально вызванными иммунодефицитами. Было показано, что удаление тимуса у новорожденного детеныша сильно ослабляет клеточные иммунные реакции, но гораздо меньше сказывается на выработке антител. У птиц можно было продемонстрировать обратный эффект, поскольку В-лимфоциты развиваются у них в фабрициевой сумке (свойственный только птицам лимфоидный орган, связанный с кишечником). Удаление фабрициевой сумки у цыплят нарушает выработку антител, но мало влияет на клеточный иммунитет. Изучение детей, родившихся с нарушенным иммунитетом, показало, что некоторые из них неспособны к выработке антител, но обладают нормальным клеточным иммунитетом, у других же наблюдается обратное соотношение. У детей с избирательным нарушением клеточных форм иммунитета почти всегда выявляются аномалии развития тимуса.
При исследовании животных с дефицитом Т-клеток (вследствие раннего удаления или повреждения тимуса) было обнаружено загадочное явление: у этих животных не только отсутствовали клеточные иммунные реакции, но была также несколько понижена способность к выработке антител. Как мы теперь знаем, это обусловлено тем, что некоторые из Т-клеток играют ключевую роль в регуляции иммунитета и действуют как помощники В-клеток в процессе гуморального ответа.
Действительно, большая часть Т-лимфоцитов играет в иммунитете регулирующую роль, усиливая или подавляя реакции других лейкоцитов. Эти клетки, называемые соответственно Т-хелперами и Т-супрессорами, объединяют в группу регуляторных клеток. Другие Т-лимфоциты, которые называют цитотоксическими Т-клетками, убивают клетки, инфицированные вирусами. Поскольку и цитотоксические Т-лимфоциты, и В-лимфоциты непосредственно участвуют в защите организма от инфекции, эти два типа лимфоцитов объединяют под названием эффекторных клеток.
18.1.3. Лимфоциты развиваются в первичных лимфоидных органах, а с чужеродными антигенами реагируют во вторичных лимфоидных органах [3]
Лимфоциты развиваются из плюрипотентных стволовых клеток, дающих начало всем клеткам крови, включая эритроциты, лейкоциты и тромбоциты (разд. 17.5.4). Эти стволовые клетки находятся главным образом в кроветворных тканях - в печени (у плода) и костном мозге (у взрослых). В-клетки у млекопитающих образуются из стволовых клеток в самих кроветворных тканях, а у птиц - в фабрициевой сумке из клеток- предшественников, которые мигрируют сюда из кроветворных тканей через кровь. Т-клетки у всех позвоночных развиваются в тимусе, куда клетки-предшественники мигрируют через кровь из кроветворных тканей. Поскольку кроветворные ткани, фабрициева сумка и тимус служат местами, где из клеток-предшественников образуются лимфоциты, их называют первичными лимфоидными органами (см. рис. 18-1). Хотя многие лимфоциты гибнут вскоре после своей дифференцировки в первичном лимфоидном органе (разд. 18.6.17), часть из них мигрирует с током крови во вторичные лимфоидные органы - главным образом в лимфатические узлы, селезенку и некоторые участки пищеварительного тракта (аппендикс, миндалины, аденоиды и пейеровы бляшки в тонком кишечнике (см. рис. 18-1). В основном именно во вторичных лимфоидных органах Т-клетки и В-клетки реагируют с чужеродными антигенами (рис. 18-3).
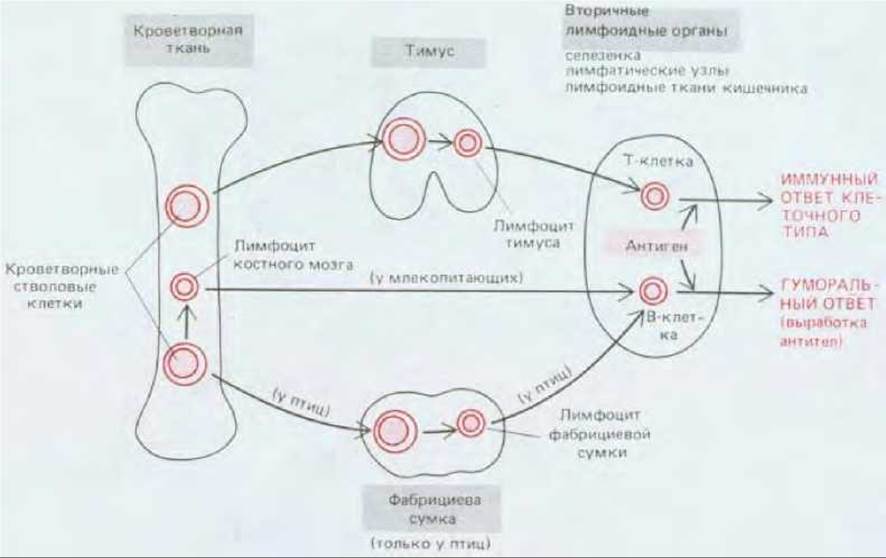
Рис. 18-3. Развитие Т- и В-лимфоцитов. И у млекопитающих, и у птиц небольшое число клеток-предшественников мигрирует с кровью в тимус, где они дифференцируются в лимфоциты тимуса. Большинство этих лимфоцитов в тимусе погибает, но некоторые мигрируют во вторичные лимфоидные органы и становятся лимфоцитами, происходящими из тимуса (Т-клетками). У птиц клетки-предшественники переходят в фабрициеву сумку, где дифференцируются в лимфоциты сумки; многие из этих лимфоцитов погибают, а некоторые мигрируют во вторичные лимфоидные органы и становятся лимфоцитами, происходящими из фабрициевой сумки (В-клетками). У млекопитающих клетки-предшественники, предназначенные для того, чтобы стать В-клетками, дифференцируются в лимфоциты в самой кроветворной ткани, а затем переходят во вторичные лимфоидные органы и становятся здесь В-клетками. Термины «Т-клетки» и «В-клетки» часто используются также для обозначения лимфоцитов тимуса и сумки (или костного мозга) соответственно. На какой стадии развития клетки-предшественники становятся детерминированными (коммитированными) к развитию в Т- или В-лимфоциты, пока не ясно. Несколько позже в этой главе мы обсудим, почему в первичных лимфоидных органах погибает так много лимфоцитов (разд. 18.4.5 и 18.6.17).
Поскольку миграция лимфоцитов из тимуса и фабрициевой сумки происходит в основном на ранних стадиях развития, удаление этих органов у взрослого животного сравнительно мало влияет на иммунный ответ; именно поэтому их роль в иммунитете так долго оставалась неизвестной. Напротив, костный мозг у млекопитающих в течение всей жизни продолжает генерировать большое число новых В-клеток (у мыши около 5-107 в сутки).
18.1.4. Маркеры клеточной поверхности позволяют различать и разделять Т- и В-клетки [4]
Т- и В-лимфоциты становятся морфологически различимыми только после стимуляции антигеном. Нестимулированные («покоящиеся») Т- и В-клетки выглядят очень сходно даже в электронном микроскопе: это небольшие - лишь немногим крупнее эритроцита-клетки, в которых большую часть объема занимает ядро (рис. 18-4, А). Те и другие активируются антигеном, вызывающим их пролиферацию и дальнейшее созревание. Активированные В-лимфоциты становятся в дальнейшем продуцентами антител. Из этих клеток наиболее зрелые-плазматические клетки с чрезвычайно развитым гранулярным эндоплазматическим ретикулом (рис. 18-4, Б). В отличие от этого активированные Т-лимфоциты содержат очень мало элементов ретикулума и не секретируют антител (рис. 18-4, В).
Поскольку и Т-, и В-лимфоциты встречаются во всех вторичных лимфоидных органах, нужно было найти методы, которые позволяли бы различать и разделять эти два типа клеток и их различные подтипы, чтобы можно было изучать их индивидуальные свойства. К счастью, различительными маркерами могут служить многочисленные гликопротеины плазматической мембраны, характерные для разных типов лимфоцитов. Например, антитела к гликопротеину Thy-І (разд. 18.6.20), который у мышей имеется на Т-, но не на В-лимфоцитах, широко используют для удаления или очистки Т-клеток из смешанной популяции лимфоцитов мыши. Антитела к гликопротеинам CD4 и CD8 (разд. 18.6.5) аналогичным образом широко используются для того, чтобы различать и разделять соответственно Т-клетки-хелперы и цитотоксические Т-клетки мыши и человека.
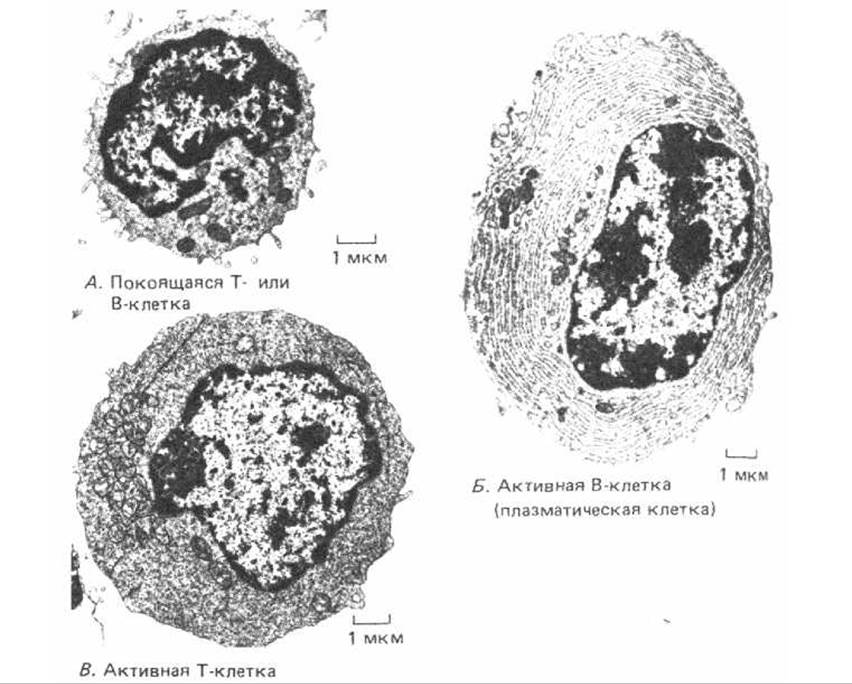
Рис. 18-4. Электронные микрофотографии покоящегося лимфоцита (А), активной В-клетки (Б) и активной Т-клетки (В). Покоящийся лимфоцит может быть Т- или В-клеткой, так как эти два вида лимфоцитов до их активации трудно различить по морфологическим признакам. Активная В-клетка (плазматическая клетка) заполнена гранулярным эндоплазматическим ретикулумом, полости которого набиты молекулами антител, тогда как в активной Т-клетке относительно мало гранулярного ретикулума, но зато очень много свободных рибосом. Все три клетки представлены при одинаковом увеличении. (С любезного разрешения: А-Dorothy Zucker-Franklin, Б - Carlo Grossi, В - Stefanello de Petris. А и Б - из D. Zucker-Franklin et al., Atlas of Blood Cells: Function and Pathology, 2nd ed. Milan, Italy: Edi. Ernies, 1988.)
18.1.5. Работа иммунной системы основана на принципе клональной селекции [5]
Самое поразительное свойство иммунной системы - то, что она может высокоспецифичным образом реагировать на миллионы чужеродны антигенов, например вырабатывая антитела, специфически взаимодействующие с тем антигеном, который вызвал их образование. Как может иммунная система обеспечивать такое разнообразие специфических антител? Одна из гипотез, весьма популярная вплоть до 40-х годов, состояла в том, что антитела синтезируются в виде развернутых полипептидных цепей, а их конечная конформация определяется антигеном, вокруг которого они сворачиваются. В то время это казалось простейшим объяснением того факта, что животные могут вырабатывать специфические антитела к молекулам, созданным человеком и не существующим в природе. Однако от такой инструктивной гипотезы пришлось отказаться, когда специалисты по химии белков установили, что трехмерная структура свернутой белковой молекулы, такой как молекула антитела, определяется только ее аминокислотной последовательностью. В самом деле, денатурированная (развернутая) молекула антитела может вновь свернуться с образованием исходного антигенсвязывающего участка даже в отсутствие антигена.

Рис. 18-5. Теория клональной селекции. Антиген активирует только те клоны Т- и В-клеток, которые уже способны на него отвечать. Предполагается, что иммунная система состоит из миллионов различных клонов лимфоцитов, из которых сотни могут быть активированы данным антигеном. В дальнейшем мы увидим, что индивидуальные клоны лимфоцитов редко отвечают на антиген автономно, как это показано на схеме и как первоначально предполагалось в теории клональной селекции. Их ответы обычно регулируются путем взаимодействий с другими клонами лимфоцитов (разд. 18.6.12). Кроме того, Т-клетки не отвечают на свободный антиген, как это показано на схеме; они отвечают только на антиген, связанный с поверхностью клетки-хозяина (разд. 18.6).
В 50-х годах инструктивная гипотеза уступила место теории клональной селекции, согласно которой каждый лимфоцит в процессе своего развития приобретает способность реагировать с определенным антигеном, еще ни разу с ним не встретившись. Это обусловлено тем, что на поверхности клетки появляются белки-рецепторы, которые специфически соответствуют какому-то антигену. Если клетка встретится с таким антигеном, то его связывание с рецепторами активирует клетку - вызовет ее размножение и созревание ее потомков. Таким образом, чужеродный антиген селективно стимулирует те клетки, у которых окажутся комплементарные ему специфические рецепторы и которые поэтому неизбежно будут реагировать именно на этот антиген - вот почему иммунные ответы антиген-специфичны (рис. 18-5).
Такую селекцию называют «клональной» потому, что иммунную систему, согласно этой теории, образуют миллионы различных клеточных «семейств», или клонов, каждый из которых состоит из Т- или В-лимфоцитов, происходящих от общего предка. Поскольку каждая клетка-предшественница уже детерминирована (или, как говорят, коммитирована) к выработке одного определенного антиген-специфического белка-рецептора, все клетки клона имеют одинаковую антигенную специфичность. Таким образом, согласно теории клональной селекции, иммунную систему по принципу ее работы можно сравнить скорее с фабрикой готовой одежды, чем с ателье, где шьют по мерке. Вопрос о том, как организм животного может вырабатывать такое множество разных антител, становится поэтому проблемой генетики, а не белковой химии.
Основные положения теории клональной селекции получили убедительные подтверждения. Например, если лимфоциты животного, которое не было иммунизировано, инкубировать в пробирке с любым из нескольких меченых антигенов, например А, В, С и D, то только очень малая доля (<0,01%) лимфоцитов будет связывать данный антиген. Это означает, что лишь немногие клетки несут специфические рецепторы для А, В, С или D. Такую интерпретацию подтверждает другой эксперимент. Антиген А делают столь высокорадиоактивным, что любая связавшая его клетка получает летальную дозу облучения; оставшаяся после этого популяция лимфоцитов уже не способна реагировать на антиген А, в то время как она продолжает нормально реагировать на В, С и D. Тот же эффект можно получить, если наполнить аффинную колонку (разд. 4.4.3) стеклянными шариками, покрытыми антигеном А, а затем пропускать через эту колонку лимфоциты. В таком опыте клетки с рецепторами для А связываются с шариками, тогда как остальные клетки проходят через колонку; клетки, прошедшие через колонку, не взаимодействуют более с А, но нормально взаимодействуют с другими антигенами (рис. 18-6). Эти два эксперимента показывают, что 1) лимфоциты коммитируются к реакции на определенный антиген еще до того, как ом подвергнутся воздействию этого антигена, и 2) коммитированные лимфоциты обладают поверхностными рецепторами, специфически связывающими данный антиген. Таким образом, подтверждаются два основных предсказания теории клональной селекции. Хотя в большинстве экспериментов такого рода изучались В-клетки и ответы, связанны с выработкой антител, другие эксперименты показали, что проявления Т-клеточного иммунитета тоже основаны на принципе клональной селекции.
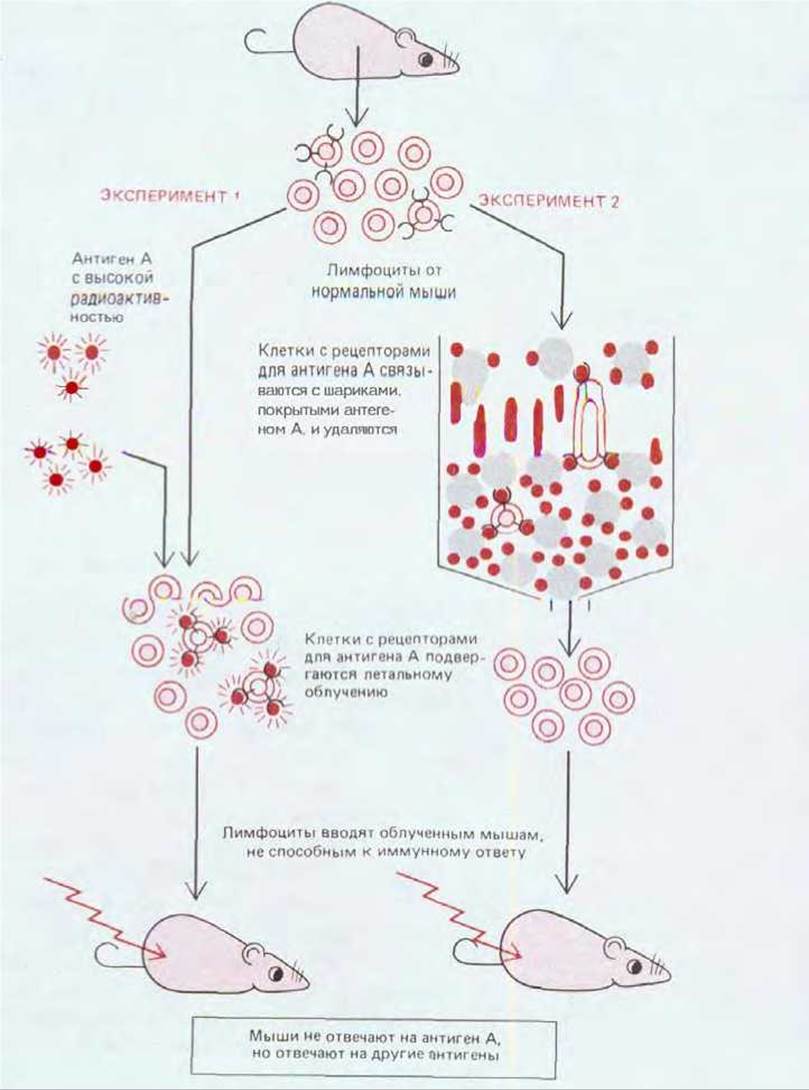
Рис. 18-6. Два типа экспериментов, подкрепляющих теорию клональной селекции. Для простоты рецепторы клеточной поверхности показаны только на лимфоцитах, способных отвечать на антиген А; на самом же деле все Т- и В-лимфоциты имеют на своей поверхности антигенспецифические рецепторы. Схематически представленные здесь эксперименты проводились главным образом с В-клетками, так как Т-клетки узнают антиген только тогда, когда он связан с поверхностью клетки-хозяина (разд. 18.6).
18.1.6. В большинстве случаев один антиген стимулирует много различных клонов лимфоцитов [6]
Большинство макромолекул, включая практически все белки и большую часть полисахаридов, могут служить антигенами. Те участки антигена, которые взаимодействуют с антиген-связывающим участком молекулы антитела или же рецептора на лимфоците, называются антигенным детерминантами (или эпитопами). Молекулы, которые хотя и присоединяются специфически к таким антиген-связывающим участкам, но не могут индуцировать иммунный ответ, называют гаптенами. Гаптены - это обычно небольшие органические молекулы; они сами по себе слишком малы для того, чтобы вызвать ответ, однако становятся полноценными антигенами, будучи присоединены к подходящей макромолекуле-носителю. Гаптены служат важными инструментами для экспериментальной иммунологии. Один из наиболее часто используемых гатенов-динитрофенильная группа (ДНФ), которую обычно пришивают к белку, чтобы сделать ее антигенной (рис. 18-7).

Рис. 18-7. Простой гаптен ДНФ, ковалентно связанный с боковой цепью лизина в белке. Гаптены могут индуцировать иммунный ответ, только будучи связаны с подобным макромолекулярным носителем.
Большинство антигенов имеет целый набор различных антигенных детерминант, стимулирующих выработку антител или Т-клеточные ответы. Некоторые детерминанты более иммуногенны (т. е. лучше индуцируют иммунитет), чем другие, и реакция на них может доминировать в общем ответе; такие детерминанты называют иммунодоминантными.
Как и следует ожидать от системы, работающей по принципу клональной селекции, даже одиночная антигенная детерминанта будет, как правило, активировать много клонов, каждый из которых будет иметь поверхностные рецепторы, обладающие своим особым, индивидуальным сродством к данной детерминанте. Например, даже сравнительно простая структура ДНФ-группы обеспечивает возможность многих различных взаимодействий, и когда эта группа связана с белком-носителем, она обычно стимулирует выработку сотен видов антител к ДНФ, каждый из которых вырабатывается отдельным клоном В-клеток. Такой ответ называют поликлональным. Когда реагирует лишь несколько клонов, ответ называют олигоклональным, а если весь ответ сводится к реакции лишь одного клона В- или Т-клеток, то его называют моноклональным. Ответы на большинство антигенов поликлональны.
Даже антиген, активирующий много клонов, воздействует лишь на ничтожную долю всей популяции лимфоцитов. Для того чтобы обеспечить встречу антигена с этими немногочисленными лимфоцитами, антигены накапливаются во вторичных лимфоидных органах, через которые непрерывно циркулируют Т- и В-лимфоциты. Антигены, проникающие в организм через пищеварительный тракт, захватываются связанными с ним лимфоидными тканями; проникающие через кожу или дыхательные пути - транспортируются с лимфой в местные лимфатические узлы; а те антигены, которые попадают в кровь, отфильтровываются в селезенке.
18.1.7. Большая часть лимфоцитов находится в непрерывной циркуляции [7]
Большинство Т- и В-лимфоцитов все время переходит из крови во вторичные лимфоидные органы и обратно. Например, в лимфатическом узле лимфоциты покидают кровяное русло, протискиваясь между специализированными эндотслиальными клетками. Пройдя через узел, они накапливаются в малых лимфатических сосудах, которые выходят из узла и соединяются с другими лимфатическими сосудами, а те проходят затем через расположенные ниже другие лимфатические узлы (рис. 18-8). Переходя во все более и более крупные сосуды, лимфоциты в конце концов попадают в главный лимфатический сосуд (грудной проток), по которому возвращаются в кровь. Такая постоянная циркуляция не только обеспечивает встречу соответствующих лимфоцитов с антигеном, но также позволяет нужным лимфоцитам встретиться друг с другом: как мы увидим, взаимодействия между специфическими лимфоцитами играют решающую роль в большинстве иммунных ответов. Циркуляция лимфоцитов зависит от специфических взаимодействий между поверхностью лимфоцита и поверхностью специализированных эндотелиальных клеток, выстилающих малые вены (посткапиллярные венулы) во вторичных лимфоидных органах: из всех клеток крови, вступающих в контакт с этими эндотелиальными клетками, только лимфоциты временно прикрепляются к ним, а затем мигрируют через посткапиллярные венулы. Моноклональные антитела (разд. 4.5.4), связываясь с поверхностью лимфоцитов и подавляя их способность присоединяться к специализированным эндотелиальным клеткам в срезах тканей, а также циркулировать in vivo, помогают определить различные «хоминг-рецепторы», от которых зависят пути миграции лимфоцитов. На поверхности большинства Т- и В-клеток имеются гликопротеины двух типов: одни для циркуляции через лимфатические узлы, а другие для циркуляции через пейеровы бляшки. Некоторые лимфоциты имеют только гликопротеин второго типа и избирательно циркулируют через пейеровы бляшки; они, в сущности, составляют специфичную для кишечного тракта подсистему лимфоцитов, специализированную для ответа на антигены, проникающие в организм через кишечник. Другие хоминг-рецепторы, по-видимому, обеспечивают сегрегацию Т- и В-клеток в отдельные участки внутри лимфоидного органа (см. рис. 18-8). Когда лимфоциты активируются антигеном, они теряют хоминг-рецепторы, опосредующие циркуляцию через лимфоидные органы, и приобретают новые рецепторы, направляющие активированные клетки к местам воспаления.

Рис. 18-8. Сильно упрощенная схема лимфатического узла человека. В-лимфоциты находятся главным образом в кортексе, где они собраны в структурах, называемых лимфатическими фолликулами. Т-лимфоциты находятся в основном в паракортикальной области. Лимфоциты обоих типов попадают в лимфатический узел из крови через небольшие специализированные вены в паракортикальной области (не показано); Т-клетки остаются в этой зоне узла, а В-клетки переходят в лимфатические фолликулы. Со временем и Т-, и В-клетки мигрируют в медуллярные синусы и покидают узел через выносящий лимфатический сосуд. Этот сосуд в конце концов вливается в кровяное русло, что позволяет лимфоцитам начать следующий цикл циркуляции через вторичный лимфоидный орган. Чужеродные антигены, попадающие в лимфатический узел, оказываются на поверхности специализированных антиген-представляющих клеток: клетки одного типа представляют антиген (в виде комплекса антиген-антитело) В-клеткам в лимфатических фолликулах, а клетки другого типа - Т-клеткам в пара-кортикальной области (разд. 18.6.10).
18.1.8. Иммунологическая память обусловлена ростом клонов и созреванием лимфоцитов [8]
Иммунная система, так же как и нервная, обладает памятью. Именно поэтому мы можем приобретать пожизненный иммунитет ко многим вирусным заболеваниям, после того как однажды подверглись воздействию вируса. Аналогичное явление можно продемонстрировать и на экспериментальных животных. Если животному однократно впрыснуть антиген А, то после лаг-периода продолжительностью в несколько дней у него появится иммунный ответ (либо антитела, либо клеточный ответ), который будет быстро (экспоненциально) усиливаться, а затем более плавно снижаться. Таково характерное протекание первичного иммунного ответа, наблюдаемого после первого контакта животного с антигеном. Если через несколько недель, месяцев или даже лет животному снова ввести антиген А, это вызовет вторичный иммунный ответ, существенно отличающийся от первичного: лаг-период будет короче, реакция сильнее и продолжительнее (рис. 18-9). Эти различия показывают, что животное «запомнило» свой первый контакт с антигеном А. Если же вместо повторной инъекции антигена А животному вводят другой антиген (например, Б), реакция в этом случае носит характер первичного, а не вторичного иммунного ответа; следовательно, вторичный ответ отражает специфическую иммунологическую память об антигене А.
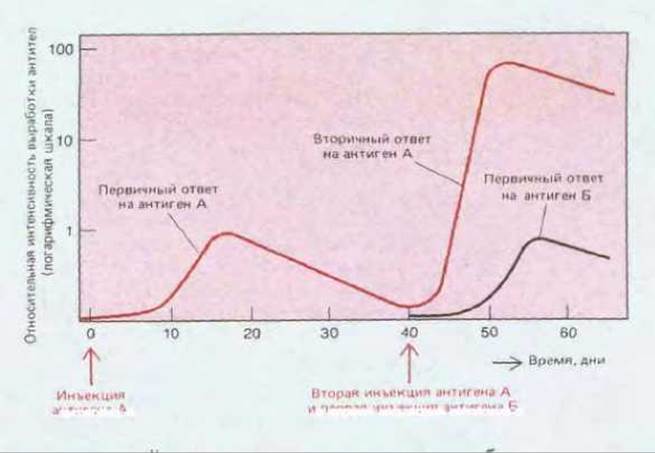
Рис. 18.9. Первичный и вторичный гуморальные ответы (образование антител), вызванные соответственно первым и вторым введением антигена А. Обратите внимание, что вторичный ответ быстрее и сильнее первичного и что он специфичен в отношении А. Это показывает, что иммунная система специфически «запомнила» ранее введенный антиген А. Такого рода иммунологическая память выявляется при изучении не только В-клеточных, но и Т-клеточных иммунных ответов
Теория клональной селекции составляет концептуальную основу для понимания клеточного механизма иммунологической памяти. Во вторичных лимфоидных органах взрослого животного популяции Т- и В-лимфоцитов одновременно содержат клетки, находящиеся по меньшей мере на трех стадиях созревания: виргильные клетки, клетки памяти и активные клетки. Когда виргильные клетки впервые встречаются с антигеном, некоторые из них стимулируются к размножению и становятся активными клетками, которые мы определяем как клетки, активно участвующие в создании иммунного ответа (активные Т-клетки реализуют клеточные ответы, а В-клетки секретируют антитела). Другие виргильные клетки вместо этого стимулируются к размножению и созреванию в клетки памяти, которые сами не дают ответа, но легко превращаются в активные клетки при последующей встрече с тем же антигеном (рис. 18-10). Как полагают, виргильные лимфоциты живут во вторичных лимфоидных органах сравнительно недолго и, вероятно, гибнут через несколько дней, если не встречаются со своим специфическим антигеном. Клетки памяти, напротив, могут жить много месяцев или даже лет без деления, постоянно циркулируя между кровью и вторичными лимфоидными органами. Кроме того, клетки памяти проявляют большую готовность отвечать на антиген, чем виргильные клетки. Позднее мы увидим (разд. 18.4.4), что повышенная готовность В-клеток памяти отвечать на антиген объясняется, в частности, более высоким сродством рецепторов этих клеток к антигену.

Рис. 18-10. Виргильные Т- или В-клетки, будучи стимулированы специфическим антигеном, делятся и созревают. Некоторые из них начинают затем давать иммунный ответ, другие же превращаются в клетки памяти. При последующей встрече с антигеном клетки памяти отвечают на него более «охотно», чем виргильные клетки: они пролиферируют и дают начало активным клеткам и новым клеткам памяти. Согласно этой модели, отдельная виргильная клетка может, в зависимости от условий, дать начало либо клетке памяти, либо активированной клетке. Согласно другой модели, здесь не представленной, клетки памяти и активированные клетки образуются из разных виргильных клеток. Которая из этих моделей верна, неизвестно.
В соответствии с этой схемой иммунологическая память создается при первичном ответе в результате того, что 1) пролиферация активированных антигеном виргильных клеток умножает число клеток памяти (экспансия клона); 2) клетки памяти имеют намного большую продолжительность жизни, чем виргильные клетки, и постоянно циркулируют между кровью и вторичными лимфоидными органами; 3) каждая клетка памяти способна более «охотно» отвечать на антиген, чем виргильная клетка. Изменения, происходящие во время первичного ответа, приводят к тому, что большая часть долгоживущих клеток в рециркулирующем пуле лимфоцитов теперь «подогнана» к антигенному окружению животного и готова к немедленному действию.
18.1.9. Отсутствие иммунного ответа на собственные антигены организма обусловлено приобретенной иммунологической толерантностью [9]
Как может иммунная система отличать «чужое» от «своего»? Одна из возможностей состоит в том, что животное наследует гены, кодирующие рецепторы для чужих, но не для собственных антигенов, и поэтому его иммунная система генетически запрограммирована таким образом, чтобы отвечать только на чужеродные антигены. Другая возможность состоит в том, что иммунная система первоначально могла быть способна отвечать и на свои, и на чужие антигены, но в раннем периоде развития могла бы «научиться» не отвечать на свои. Было показано, что верна вторая из этих гипотез. Первым свидетельством в пользу этого явилось наблюдение, сделанное в 1945 г. Как правило, при пересадке ткани от одного индивидуума другому трансплантат распознается иммунной системой как чужеродный и отторгается. Оказалось, однако, что этого не происходит при пересадках кожи между дизиготными (развившимися из двух оплодотворенных яйцеклеток, т. е. неидентичными) коровами-близнецами, которые во время внутриутробного развития могли обмениваться клетками крови вследствие спонтанного сращения их плацент. Эти результаты позднее были воспроизведены на курах (путем соединения кровеносных сосудов двух разных эмбрионов) и на мышах (путем введения новорожденным мышатам клеток селезенки от мышей другой линии - эти клетки выживали в течение большей части жизни мыши-реципиента). В обоих случаях, когда животные становились взрослыми, можно было пересаживать им ткань от временно присоединенной особи или от особи-донора, и трансплантат приживался (рис. 18-11), тогда как ткани, пересаженные от других, контрольных животных, отторгались. Таким образом, постоянное присутствие чужих антигенов начиная с того времени, когда иммунная система еще к созрела, приводит к долговременной ареактивности по отношению к этим антигенам. Такое состояние индуцированной антиген-специфической неспособности к иммунному ответу получило название приобретенной иммунологической толерантности.
Есть убедительные данные в пользу того, что неспособность иммунной системы животного реагировать на свои собственные макромолекулы (естественная иммунологическая толерантность) приобретаете тем же самым путем - она не врожденная. Например, нормальные мыши не дают иммунного ответа на свой собственный белок крови - компонент комплемента С5 (разд. 18.5.1). Однако мутантные мыши, у которых нет кодирующего С5 гена (в остальном генетически идентичные нормальным мышам), могут давать иммунную реакцию на этот белок Таким образом, ясно, что иммунная система потенциально способна реагировать на антигены собственного организма, но «обучается» не делать этого. По крайней мере в некоторых случаях процесс «обучения» включает элиминацию лимфоцитов, реагирующих на «свое» (разд. 18.6.17), но мы не знаем, как она происходит. Полагают, что многие из таких лимфоцитов элиминируются в первичных лимфоидных органах при встрече с антигеном. Подобный отрицательный ответ на антиген мог бы быть обусловлен специфическим окружением в этих органах или исключительной активностью новообразованных лимфоцитов. Возможно, из-за того, что новые реагирующие на «свое» лимфоциты продолжают вырабатываться из стволовых клеток в течение всей жизни, для поддержания естественной толерантности требуется постоянное присутствие «своих» антигенов. Если такой антиген, как С5, удалить, организм животного спустя недели или месяцы снова приобретает способность отвечать на него иммунной реакцией.

Рис. 18-11. Показанный здесь кожный лоскут, трансплантированный взрослой белой мыши от взрослой коричневой мыши, выжил в течение многих недель только потому, что белую мышь сделали иммунологически толерантной, введя ей сразу после рождения клетки крови от коричневой мыши. (С любезного разрешения Leslie Brent, из I. Roitt, Essential Immunology, 6th ed. Oxford, U.K.: Blackwell Scientific, 1988.)
Толерантность к собственным антигенам иногда нарушается, что приводит к реакции Т- или В-клеток (или тех и других) на тканевые антигены собственного организма. Примером таких аутоиммунных заболеваний может служить миастения (myasthenia gravis). При этой болезни образуются антитела к ацетилхолиновым рецепторам на волокнах скелетных мышц (разд. 6.4.18); эти антитела препятствуют нормальному функционированию рецепторов. Такие больные страдают мышечной слабостью и могут умереть из-за недостаточной функции дыхательной мускулатуры.
18.1.10. Иммунологическую толерантность к чужеродным антигенам можно индуцировать и у взрослых животных [10]
У взрослого животного создать иммунологическую толерантность к чужеродным антигенам, как правило, гораздо труднее, чем на ранней стадии развития. Но в отношении некоторых антигенов это можно сделать экспериментально, вводя антиген 1) в очень больших дозах, 2) многократно в очень малых дозах, 3) вместе с иммунодепрессантом или 4) внутривенно после химической сшивки антигена с поверхностью В-лимфоцитов или ультрацентрифугирования антигена с целью удалить все агрегаты (это делает неэффективными нормальные механизмы презентации антигена, см. разд. 18.6.10). Таким образом, связывание антигена с комплементарными ему рецепторами на Т- или В-лимфоцитах может либо стимулировать лимфоцит к делению и созреванию, в результате чего он превращается в активную клетку или клетку памяти, либо элиминировать или инактивировать лимфоцит, что приводит к толерантности. Молекулярные механизмы, определяющие исход дела, изучены недостаточно. Вызовет ли антиген активацию или индуцирует толерантность, зависит главным образом 1) от степени зрелости лимфоцита, 2) от природы и концентрации антигена и 3) от сложных взаимодействий между лимфоцитами разных классов и между лимфоцитами и специализированными антиген-представляющими клетками, которые будут описаны позже.
Заключение
Иммунная система вырабатывалась в процессе эволюции позвоночных для защиты от инфекций. Она состоит из миллионов клонов лимфоцитов. Лимфоциты каждого клона несут на своей поверхности рецептор, позволяющий им связывать ту или иную «антигенную детерминанту»-определенную группировку атомов в молекуле антигена. Существуют два класса лимфоцитов: В-клетки, вырабатывающие антитела, и Т-клетки, которые осуществляют иммунные реакции клеточного типа.
Уже на ранних стадиях своего развития многие лимфоциты, реагирующие с антигенными детерминантами макромолекул собственного организма, элиминируются или инактивируются; в результате иммунная система в норме реагирует только на чужеродные антигены. Связывание чужеродного антигена с лимфоцитом вызывает иммунный ответ, направленный против этого антигена. При этом некоторые из лимфоцитов размножаются и созревают, превращаясь в долгоживущие клетки памяти, так что при вторичной встрече организма с тем же антигеном иммунный ответ развивается быстрее и оказывается более сильным.