Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Половые клетки и оплодотворение
Мейоз
Понимание того факта, что половые клетки гаплоидны и поэтому должны формироваться с помощью особого механизма клеточного деления, пришло в результате наблюдений, которые к тому же едва ли не впервые навели на мысль, что хромосомы содержат генетическую информацию. В 1883 г. было обнаружено, что ядра яйца и спермия определенного вида червей содержат лишь по две хромосомы, в то время как в оплодотворенном яйце их уже четыре. Хромосомная теория наследственности могла, таким образом, объяснить давний парадокс, состоящий в том, что роль отца и матери в определении признаков потомства часто кажется одинаковой, несмотря на огромную разницу в размерах яйцеклетки и сперматозоида.
Еще один важный смысл упомянутого открытия состоял в том, что половые клетки должны формироваться в результате ядерного деления особого типа, при котором весь набор хромосом делится точно пополам. Деление такого типа носит название мейоз (слово греческого происхождения, означающее «уменьшение». Название другого вида деления клеток-митоз-происходит от греческого слова mitos, означающего «нить»; в основе такого выбора названия лежит нитеподобный вид хромосом при их конденсации во время деления ядра - данный процесс имеет место как при обычном, так и при мейотическом делении.) Поведение хромосом во время мейоза, когда происходит редукция их числа, оказалось более сложным, чем предполагали раньше. Поэтому важнейшие особенности мейотического деления удалось установить только к началу 30-х годов в итоге огромного числа тщательных исследований, объединивших цитологию и генетику.
15.2.1. При мейозе происходит не одно, а два деления ядра
Диплоидные ядра содержат по две копии каждой хромосомы (это не относится лишь к половым хромосомам), одна из которых происходит от мужского родителя, а другая - от женского. Эти две копии называются гомологами, и в большинстве клеток они ведут себя как совершенно независимые хромосомы. Когда благодаря репликации ДНК каждая хромосома удваивается, две ее копии остаются сначала соединенными вместе (их называют сестринскими хроматидами). При обычном клеточном делении (описанном в гл. 13) сестринские хроматиды выстраиваются в экваториальной плоскости веретена таким образом, что их кинетохорные волокна направлены к противоположным полюсам. В результате сестринские хроматиды в анафазе отделяются друг от друга (теперь они называются хромосомами), и каждая дочерняя клетка наследует по одной копии каждого гомолога (см. разд. 13.5). Между тем гаплоидные гаметы, образовавшиеся при делении диплоидной клетки путем мейоза, содержат по одной хромосоме каждой гомологичной пары (отцовского или материнского происхождения), т. е. только половину исходного числа хромосом. В связи с этим к аппарату клеточного деления здесь предъявляется дополнительное требование: гомологи должны иметь возможность «узнавать» друг друга и соединяться в пары, перед тем как они выстроятся на экваторе веретена. Такое спаривание, или конъюгация, гомологичных хромосом материнского и отцовского происхождения происходит только в мейозе (рис. 15-8); подробности этого процесса будут рассмотрены позже.
При наличии механизма конъюгации отцовских и материнских гомологичных хромосом и их последующего расхождения мейоз мог бы в принципе осуществляться путем видоизменения одного митотического цикла, если бы в нем выпала фаза удвоения хромосом (S) и гомологи спаривались перед фазой М. Тогда в результате следующего клеточного деления могли бы непосредственно образоваться две гаплоидные клетки. Однако на самом деле процесс мейоза более сложен. Перед конъюгацией каждый из гомологов подвергается удвоению, образуя пару тесно связанных сестринских хроматид аналогично тому, как это происходит при обычном клеточном делении. Специфические особенности мейоза проявляются лишь после завершения репликации ДНК. Вместо того чтобы отделиться друг от друга, сестринские хроматиды ведут себя как единое целое (как будто дупликация хромосом не произошла): каждый дуплицированный гомолог конъюгирует с партнером, образуя структуру, состоящую из четырех хроматид и называемую бивалентом. Бивалент располагается на экваторе веретена, и в анафазе дуплицированные гомологи (каждый из которых состоит из двух сестринских хроматид) отделяются друг от друга и расходятся к противоположным полюсам причем в каждом из них две сестринские хроматиды остаются соединенными. Таким образом, при первом делении мейоза каждая дочерняя клетка наследует две копии одного из двух гомологов и поэтому содержит диплоидное количество ДНК. Однако она отличается от обычных диплоидных клеток в двух отношениях: 1) обе копии ДНК каждой хромосомы происходят лишь от одной из двух гомологичных хромосом, имевшихся в исходной клетке (хотя, как мы увидим, в результате генетической рекомбинации происходит некоторое перемешивание материнских и отцовских ДНК), и 2) эти две копии клетка получает в виде тесно связанных сестринских хроматид, составляющих единую хромосому (рис. 15-8).
Теперь образование гаплоидных ядер гамет может очень просто происходить в результате второго деления мейоза, при котором хромосомы выстраиваются на экваторе нового веретена и без дальнейшей репликации ДНК сестринские хроматиды отделяются друг от друга, как при обычном митозе, образуя клетки с гаплоидным набором ДНК. Таким образом, мейоз состоит из двух клеточных делений, следующих за единственной фазой удвоения хромосом, так что из каждой клетки, вступающей в мейоз, получаются в итоге четыре гаплоидные клетки (рис. 15-8). Иногда процесс мейоза протекает аномально, и гомологи не могут отделиться друг от друга - это явление называется нерасхождением хромосом. Некоторые из образующихся в этом случае гаплоидных клеток получают недостаточное количество хромосом, в то время как другие приобретают их лишние копии. Из подобных гамет формируются неполноценные эмбрионы, большая часть которых погибает.

Рис. 15-8. Сравнение мейоза с обычным митозом (схема). Для простоты показана только одна пара гомологичных хромосом. Спаривание гомологичных хромосом происходит только в мейозе; поскольку перед спариванием каждая хромосома удваивается и состоит из двух сестринских хроматид, для образования гаплоидных гамет необходимы два клеточных деления. Поэтому из любой диплоидной клетки, вступающей в мейоз, образуются четыре гаплоидные клетки. В мейозе при конъюгации гомологичных хромосом между ними осуществляется кроссинговер; объяснение сути этого явления дается позже.

Рис. 15-9. Схема, иллюстрирующая два основных механизма перераспределения генетического материала во время мейоза. А. У организма с п хромосомами в результате независимого расхождения гомологичных хромосом в первом делении мейоза может получиться 2" различных гаплоидных гамет. В данном случае n = 3 и может быть 8 различных типов гамет. Б. В I профазе мейоза происходит кроссинговер-гомологичные хромосомы обмениваются участками, что ведет к перераспределению генов. В последовательностях ДНК двух гомологов всегда имеется большое число незначительных различий, поэтому оба механизма увеличивают наследственную изменчивость организмов, размножающихся половым путем.
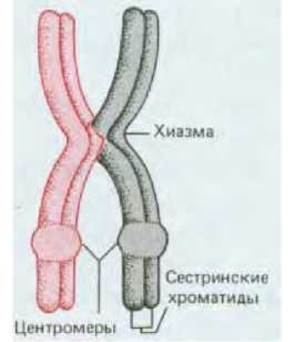
Рис. 15-10. Схематическое изображение спаренных гомологичных хромосом при переходе к I метафазе мейоза. В предшествующей профазе произошел один кроссинговер, и в результате образовалась одна хиазма. Обратите внимание, что четыре хроматиды сгруппированы в две пары сестринских нитей, причем в каждой паре они тесно сближены не только в области центромеры, но и по всей длине. Поэтому всю такую группу хроматид часто называют бивалентом.
15.2.2 Пересортировка генов усиливается благодаря кроссинговеру между гомологичными несестринскими хроматидами
Как мы уже видели, гены могут перемешиваться благодаря слиянию гамет двух различных особей. Однако генетические изменения осуществляются не только этим путем. Никакие два потомка одних и тех же родителей (если только это не идентичные близнецы) не будут абсолютно одинаковыми. Дело в том, что задолго до слияния двух гамет, во время мейоза, осуществляются два различных вида пересортировки генов.
Один вид пересортировки - это результат случайного распределения разных материнских и отцовских гомологов между дочерними клетками при 1-м делении мейоза; каждая гамета получает свою, отличную от других выборку материнских и отцовских хромосом (рис. 15-9 А). Из одного только этого факта следует, что клетки любой особи могут в принципе образовать 2" генетически различающихся гамет, где n-гаплоидное число хромосом. Например, у человека каждый индивидуум способен образовать по меньшей мере 223 = 8,4-106 генетически различных гамет. Однако на самом деле число возможных гамет неизмеримо больше из-за кроссинговера (перекреста) - процесса, происходящего во время длительной профазы 1-го деления мейоза, когда гомологичные хромосомы обмениваются участками. У человека в каждой паре гомологичных хромосом кроссинговер происходит в среднем в двух-трех точках. Как показано на рис. 15-9 Б, такой процесс «перетасовывает» гены любой хромосомы в гаметах.
При кроссинговере происходит разрыв двойной спирали ДНК в одной материнской и одной отцовской хроматиде, а затем получившиеся отрезки воссоединяются «наперекрест» (процесс генетической рекомбинации). То, что известно о деталях молекулярного механизма этого процесса, в общих чертах представлено в гл. 5. Рекомбинация происходит в профазе 1-го деления мейоза, когда две сестринские хроматиды так тесно сближены друг с другом, что их невозможно увидеть в отдельности (см. ниже). Гораздо позже в этой растянутой профазе становятся ясноразличимы две отдельные хроматиды каждой хромосомы. В это время видно, что они связаны своими центромерами и тесно сближены друг с другом по всей длине. Два гомолога остаются связанными в тех точках, где произошел кроссинговер между отцовской и материнской хроматидами. Видно, что в каждой такой точке, которую называют хиазмой, две из четырех хроматид перекрещиваются (рис. 1510). Таким образом, хиазмы - это морфологический результат произошедшего кроссинговера, который сам по себе недоступен для наблюдения.
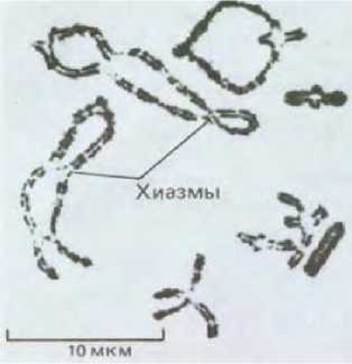
Рис. 15-11. Световая микрофотография бивалентов с множественными хиазмами на стадии диплотены. Изображенные здесь крупные хромосомы прямокрылого особенно удобный объект для цитологических исследований. (С любезного разрешения Bernard John.)
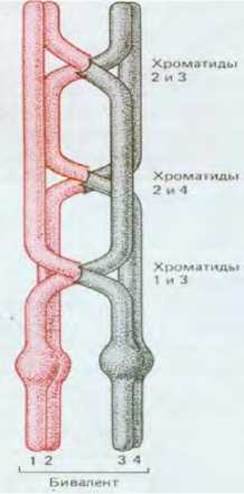
Рис. 15-12. Представлены три хиазмы - результат трех отдельных перекрестов. Каждая из двух хроматид одной хромосомы может перекрещиваться с любой хроматидой другой хромосомы бивалента. Например, в данном случае хроматида 3 обменялась участками одновременно с двумя хроматидами - 1 и 2.
На этой стадии мейоза гомологи в каждой паре (или биваленте) остаются связанными друг с другом по меньшей мере одной хиазмой. Во многих бивалентах бывает большее число хиазм, так как возможны множественные перекресты между гомологами (рис. 15-11 и 15-12).
15.2.3. В конъюгации хромосом участвует синаптонемальный комплекс
В профазе 1-го деления мейоза во время конъюгации (синопсиса) и разделения хромосом в них происходят сложнейшие морфологические изменения. В соответствии с этими изменениями профаза делится на пять последовательных стадий - лептотену, зиготену, пахитену, диплотену и диакинез (рис. 15-13). Самое поразительное явление - это инициация тесного сближения хромосом в зиготене, когда между парами сестринских хроматид в каждом биваленте начинает формироваться специализированная структура, называемая синаптонемальным комплексом. Момент полной конъюгации хромосом считают началом пахитены, которая обычно продолжается несколько дней; после разделения хромосом наступает стадия диплотены, когда впервые становятся видны хиазмы.
Для генетической рекомбинации необходимо тесное сближение рекомбинирующих хромосом. Синаптонемальный комплекс, который формируется перед самой пахитеной и распадается сразу после нее, удерживает гомологичные хромосомы рядом, скрепляя их по всей длине; полагают, что он необходим для осуществления кроссинговера. Синаптонемальный комплекс представляет собой длинное белковое образование, напоминающее веревочную лестницу, к противоположным сторонам которого плотно прилегают два гомолога (бивалент, рис. 15-14). Сестринские хроматиды каждого гомолога остаются тесно сближенными, а их ДНК образует многочисленные петли по ту же сторону от белковой «лестницы». Таким образом, хотя гомологичные хромосомы в синаптонемальном комплексе сближены по всей длине, материнские и отцовские хроматиды, которые впоследствии будут обмениваться участками, остаются по разные стороны от «лестницы», причем разделяющее их расстояние превышает 100 нм.
Как показывают цитологические исследования, конъюгации хромосом предшествует формирование белковой нити вдоль каждого из гомологов. По мере осуществления конъюгации эти нити, по-видимому, сближаются, превращаясь в боковые элементы синаптонемального комплекса и образуя две стороны белковой «лестницы». И первоначальные нити, и эти боковые элементы содержат белок, который очень хорошо окрашивается серебром, что позволяет видеть эти структуры как с помощью светового микроскопа, так и на электронных микрофотографиях (рис. 15-15). Мы не знаем, что заставляет гомологичные участки хромосом точно ориентироваться друг против друга на стадии зиготены. Вряд ли для этого необходимо спаривание комплементарных оснований по всей длине взаимодействующих хромосом, поскольку хроматин одного из гомологов в синаптонемальном комплексе расположен достаточно далеко от хроматина другого гомолога, а в отдельных случаях синаптонемальный комплекс может объединять участки двух негомологичных хромосом. Одно из возможных объяснений состоит в том, что конъюгация хромосом начинается с взаимодействия комплементарных пар оснований ДНК в небольших специфических участках каждой хромосомы. Затем синаптонемальный комплекс соединяет оставшиеся области ориентированных друг против друга хромосом. Какой-то механизм такого рода локального соответствия необходим для объяснения того факта, что наличие в одной из двух гомологичных хромосом инвертированного участка обычно (хотя и не всегда) приводит к местному нарушению нормального синапсиса во время зиготены, так что гомологичные гены могут конъюгировать даже в области инверсии (рис. 15-16 и 15-17). Отдельные стадии мейоза показаны на рис. 15-18, где дается и подробное описание соответствующих процессов.

Рис. 15-13. Последовательность событий при синапсисе и разъединении хромосом в I профазе мейоза. Полностью сформированный синаптонемальный комплекс существует на протяжении всей стадии пахитены.
15.2.4. Как полагают, обмены между хроматидами происходят при участии рекомбинационных узелков
Синаптонемальный комплекс обеспечивает структурную основу, необходимую для рекомбинационных событий, но сам он, вероятно, непосредственно в них не участвует. Как полагают, важную роль в этих событиях играют рекомбинационные узелки, которые представляют собой очень крупные белковые комплексы с диаметром около 90 нм (для сравнения заметим, что крупная молекула глобулярного белка массой 400000 дальтон имеет диаметр порядка 10 нм). Рекомбинационные узелки «сидят» на некоторых расстояниях друг от друга на «лестнице» синаптонемального комплекса, между двумя гомологичными хроматидами (см. рис. 15-14). Предполагается, что это место расположения крупных мультиферментных «рекомбинационных аппаратов», которые подтягивают друг к другу локальные участки ДНК материнской и отцовской хроматид через область синаптонемального комплекса шириной 100 нм.
О такой функции рекомбинационных узелков говорят некоторые косвенные данные:
1. Общее число узелков примерно равно общему числу хиазм, наблюдаемых позже в профазе.
2. Узелки распределены вдоль синаптонемального комплекса таким же образом, как и перекресты; например, подобно перекрестам, узелки отсутствуют в тех областях, где синаптонемальный комплекс соединяет отрезки гетерохроматина. Кроме того, генетические и цитологические исследования показывают, что произошедший кроссинговер препятствует осуществлению другого кроссинговера в близлежащем участке хромосомы. Точно так же и узелки, как правило, не располагаются очень близко друг к другу.
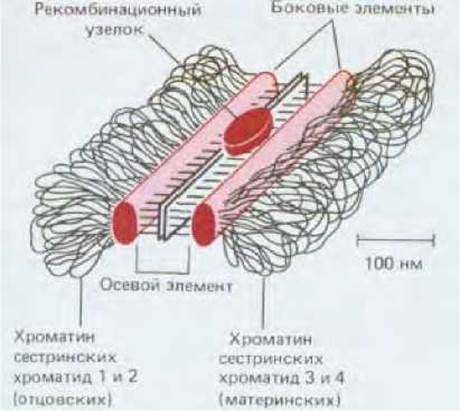
Рис. 15-14. Схематическое изображение типичного синаптонемального комплекса. Здесь представлен лишь небольшой участок этого длинного, похожего на лестницу образования. Показаны боковые и осевой (центральный) элементы комплекса, а также рекомбинационный узелок. Сходные синаптонемальные комплексы обнаруживаются у самых различных организмов - от дрожжей до человека. Однако об образующих эти комплексы белковых молекулах почти ничего не известно.
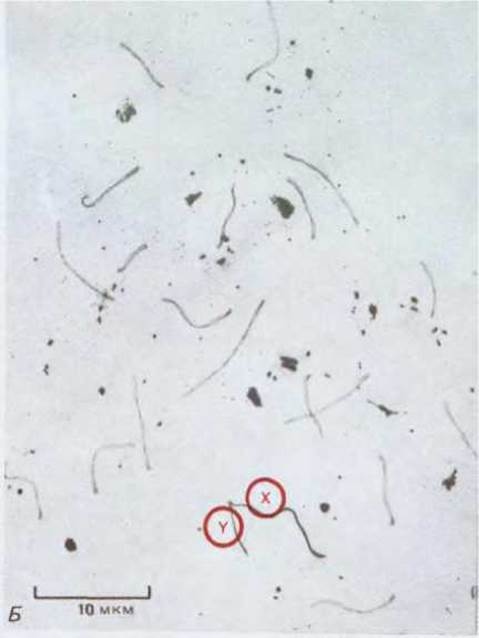

Рис. 15-15. Электронные микрофотографии полных наборов хромосом в окрашенных серебром давленых препаратах сперматоцитов мыши на (А) ранней (зиготена), (Б) средней (пахитена) и (В) поздней (диплотена) стадиях профазы первого деления мейоза. (Фото любезно предоставлены Montrose J. Moses.)
А. Спаривание (зиготена). Перед конъюгацией хромосом белковые нити отделены друг от друга; затем они сближаются и, когда в одном или нескольких местах инициации синапсиса между хромосомами установится надлежащее расстояние, начинает формироваться синаптонемальный комплекс (часто с конца хромосомы). На примере половых хромосом (X и Y) видно, что для спаривания нередко приходится преодолевать огромные расстояния внутри ядра, однако механизм этого движения неизвестен. Темные тельца-ядрышки.
Б. Завершение спаривания (пахитена). Синапсис завершается, когда синаптонемальные комплексы связывают попарно все гомологичные аутосомы. Х- и Y-хромосомы конъюгируют не полностью. Происходит кроссинговер между хроматидами, которые на таких фотографиях неразличимы.
В. Расхождение хромосом (диплотена). Перед разрушением белковых нитей последние отделяются друг от друга, что означает окончание синапсиса. На препарате они еще кое-где связаны сохранившимися отрезками синаптонемального комплекса; как полагают, это те места, в которых произошел кроссинговер. Позже, когда хроматин конденсируется и становятся различимыми хроматиды, на места кроссинговера указывают хиазмы.
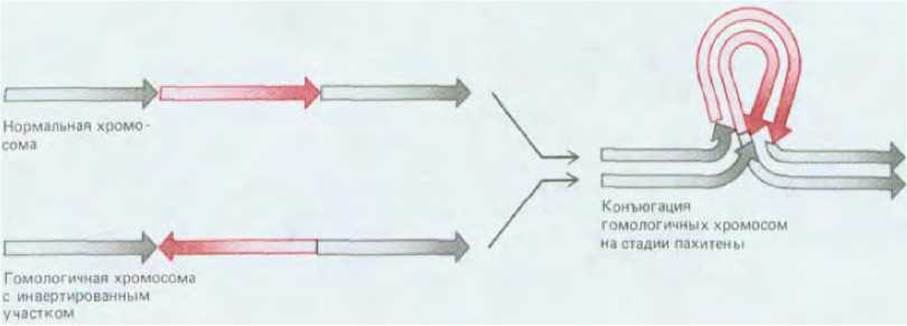
Рис. 15-16. Схема образования синаптонемального комплекса между нормальной хромосомой и ее гомологом, имеющим инвертированный участок.
Подобные структуры указывают на то, что гомологичные хромосомы конъюгируют благодаря локальному сходству определенных участков.
Сравните данный рисунок с рис. 15-17.
3. Некоторые мутации у дрозофилы приводят к аномальному распределению перекрестов по длине хромосом и резко пониженной частоте рекомбинаций; при этом у мух оказывается меньше рекомбинационных узелков и их размещение вдоль хромосомы изменено так же, как и распределение перекрестов. Такая корреляция служит веским доводом в пользу того, что каждый кроссинговер определяется локализацией одного узелка.
4. Как полагают, при генетической рекомбинации в области каждого кроссинговера происходит синтез некоторого количества ДНК (см. разд. 5.4.5). Метод радиоавтографии в сочетании с электронной микроскопией позволяет показать, что радиоактивные предшественники включаются в пахитенную ДНК главным образом в области рекомбинационных узелков или поблизости от них.
Поскольку рекомбинационных узелков бывает примерно столько же, сколько происходит перекрестов, можно думать, что эти узелки очень эффективно вызывают рекомбинацию между хроматидами двух гомологичных хромосом. К сожалению, о структуре рекомбинационных узелков и механизме их действия пока ничего не известно.
15.2.5. Хиазмы играют важную роль в расхождении хромосом во время мейоза
Кроссинговер не только способствует перетасовке генов, но играет также важнейшую роль при расхождении двух гомологов в дочерние ядра. Дело в том, что именно хиазмы удерживают вместе материнские и отцовские гомологи до анафазы I, выполняя здесь ту же функцию и центромеры в обычном митозе. У мутантных организмов с пониженной частотой кроссинговера в мейозе у отдельных пар хромосом отсутствуют хиазмы, и такие хромосомы не способны нормально расходиться. В результате значительная доля образующихся гамет содержит слишком много или слишком мало хромосом - последний случай служит примером не расхождения хромосом.
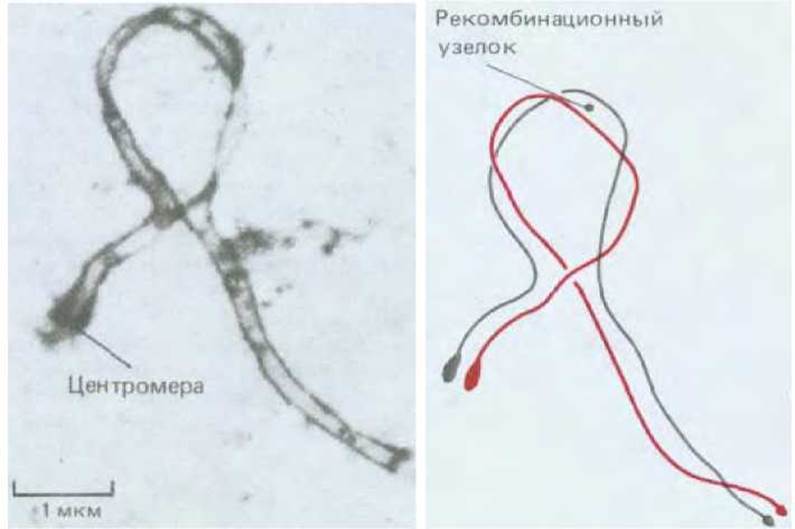
Рис. 15-17. Тесная конъюгация двух гомологичных хромосом мыши, одна из которых содержит инверсию, на стадии пахитены. В петле виден рекомбинационный узелок (см. разд. 15.2.4). Слева - электронная микрофотография; справа поясняющая схема. (P. A. Poorman et al., Chromosoma, 83, 419. 1981.)

Рис. 15-18. Схемы, показывающие видимые изменения двух гомологичных хромосом на протяжении мейоза. Представлен ход событий в клетках млекопитающих, хотя весьма сходную картину можно наблюдать и в клетках многих других организмов. А - пять стадий первой профазы мейоза (А); Б - последующие стадии мейоза.
ЛЕПТОТЕНА. Профаза I начинается со стадии лептотены, когда видно, что каждая хромосома, изменив свою интерфазную конформацию, переходит в конденсированную форму, образуя длинное, тонкое волокно с белковой осевой нитью. Каждая хромосома обоими концами прикреплена к ядерной мембране с помощью специализированной структуры, называемой прикрепительным диском. Хотя каждая хромосома уже реплицировалась и состоит из двух сестринских хроматид, эти хроматиды очень тесно сближены, и поэтому каждая хромосома кажется одиночной (отдельные хроматиды не различимы вплоть до поздней профазы - до стадии диплотены или диакинеза)
ЗИГОТЕНА. Моментом перехода лептотены в зиготену считают начало синапсиса - тесной конъюгации двух гомологов. Конъюгация часто начинается с того, что гомологичные концы двух хромосом сближаются на ядерной мембране, а затем процесс соединения гомологов распространяется вдоль хромосом от обоих концов. В других случаях синапсис может начинаться во внутренних участках хромосом и продолжаться по направлению к их концам с тем же конечным результатом. Как полагают, каждый ген приходит в соприкосновение с гомологичным ему геном другой хромосомы. Когда гомологи конъюгируют, их белковые нити сближаются, образуя два боковых элемента длинного образования, напоминающего лестницу и называемого синаптонемальным комплексом. Каждую пару хромосом, образовавшуюся в I профазе мейоза, обычно называют бивалентом; но, поскольку каждая гомологичная хромосома пары состоит из двух тесно сближенных сестринских хроматид, для каждой пары больше подходит другое название тетрада
ПАХИТЕНА. Как только завершается синапсис по всей длине хромосом, клетки вступают в стадию пахитены, на которой они могут оставаться несколько суток. На этой стадии в продольной щели синаптонемального комплекса появляются крупные рекомбинационные узелки, которым приписывают важную роль в обмене участками между хромосомами. Такие обмены приводят к перекрестам между двумя несестринскими хроматидами: в обменах участвует по одной хроматиде из двух спаренных хромосом. В пахитене перекресты еще не видны, но позднее все они проявляются в виде хиазм
ДИПЛОТЕНА. Стадия диплотены в 1 профазе мейоза начинается с разделения конъюгировавших хромосом. Синаптонемальный комплекс распадается, что позволяет двум гомологичным хромосомам бивалента несколько отодвинуться друг от друга. Однако они все еще связаны одной или несколькими хиазмами, т. е. местами, где произошел кроссинговер. В ооцитах (развивающихся яйцеклетках) диплотена может растянуться на месяцы или годы, так как именно на этой стадии хромосомы конденсируются и синтезируют РНК, обеспечивая яйцеклетку резервными веществами. В особых случаях диплотенные хромосомы становятся исключительно активными в отношении синтеза РНК; такие хромосомы типа ламповых щеток находят у амфибий и некоторых других организмов ДИАКИНЕЗ. Диплотена незаметно переходит в диакинез - стадию, предшествующую метафазе, когда прекращается синтез РНК и хромосомы конденсируются, утолщаются и отделяются от ядерной мембраны. Теперь ясно видно, что каждый бивалент содержит четыре отдельные хроматиды, причем каждая пара сестринских хроматид соединена центромерой, тогда как несестринские хроматиды, претерпевшие кроссинговер, связаны хиазмами. После окончания длительной профазы I два ядерных деления без разделяющего их периода синтеза ДНК доводят процесс мейоза до конца. Эти стадии обычно занимают не более 10% всего времени, необходимого для мейоза, и они носят те же названия, что и соответствующие стадии митоза. В оставшейся части первого деления мейоза различают метафазу I, анафазу I и телофазу I. К концу первого деления хромосомный набор редуцируется, превращаясь из тетраплоидного в диплоидный, совсем как при митозе, и из одной клетки образуются две. Решающее различие состоит в том, что при первом делении мейоза в каждую клетку попадают две сестринские хроматиды соединенные в области центромеры, а при митозе - две разделившиеся хроматиды. Далее, после кратковременной интерфазы II, в которой хромосомы не удваиваются, быстро происходит второе деление - профаза II, анафаза II и телофаза II. В результате из каждой диплоидной клетки, вступившей в мейоз, образуются четыре гаплоидных ядра.
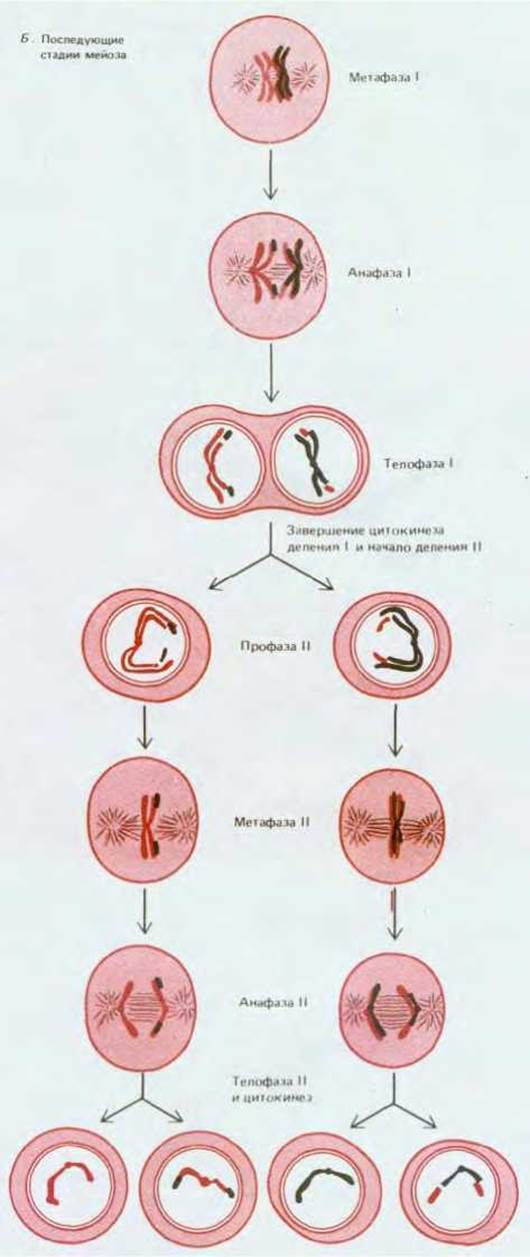

Рис. 15-19. Сравнение механизмов упорядоченного расположения хромосом в метафазе и их расхождения в анафазе при первом и втором делениях мейоза. Во втором делении используются те же механизмы, что и в обычном митозе (см. гл. 13).
Существуют по меньшей мере два важных различия между механизмами расхождения хромосом в обычном митозе и при первом делении мейоза:
1) если при митозе нити веретена, прикрепленные к кинетохорам двух сестринских хроматид, отходят в противоположных направлениях, то в первой метафазе мейоза эти нити у обеих сестринских хроматид отходят в одном и том же направлении (рис. 15-19);
2) при митозе расхождение хроматид к полюсам инициируется отделением друг от друга сестринских кинетохоров (так начинается анафаза, разд. 13.5.7), тогда как в первой анафазе мейоза это движение, по-видимому, начинается в результате исчезновения мало изученных сил, которые удерживают плечи сестринских хроматид в тесно сближенном состоянии, что в свою очередь ведет к распаду хиазм, связывающих материнские и отцовские хромосомы (см. рис. 15-9). Это не только позволяет объяснить тот факт, что у многих организмов хиазмы необходимы для нормального расположения хромосом в метафазе I, но и дает ответ на вопрос, почему у хромосом, образующихся в анафазе I, плечи сестринских хроматид обыкновенно не слипаются, что придает им необычный «развернутый» вид и делает их похожими на митотические хромосомы (рис. 15-19).
15.2.6. Расхождение половых хромосом тоже обеспечивается их конъюгацией
Мы объяснили, каким образом конъюгация гомологичных хромосом обусловливает их расхождение в две дочерние клетки. Но как обстоит дело с половыми хромосомами, которые у самцов млекопитающих не гомологичны? У самок имеются две Х-хромосомы, которые конъюгируют и расходятся так же, как другие гомологи. Однако самцы обладают одной Х - и одной Y-хромосомой, и эти хромосомы должны конъюгировать во время первой метафазы, чтобы сперматозоиды содержали либо Х -, либо Y-хромосому, но исключалось наличие или отсутствие в них сразу обеих половых хромосом. Требуемая конъюгация становится возможной благодаря наличию небольшого участка на конце каждой из этих хромосом, где между ними имеется гомология; эта гомология позволяет половым хромосомам X и Y спариваться во время первой профазы мейоза (рис. 15-15, Б). Таким образом, гарантируется конъюгация Х- и Y-хромосом и образование спермиев только двух типов: одни содержат Y-хромосому и дают начало эмбриону мужского пола, а другие - Х-хромосому и дают начало эмбриону женского пола.
15.2.7. Второе деление мейоза напоминает обычный митоз
Мейоз состоит из двух последовательных клеточных делений, первое из которых длится почти столько же, сколько весь мейоз, и гораздо сложнее второго (рис. 15-20). Первое деление отличается рядом уникальных особенностей. Например, репликация ДНК во время подготовительной фазы S, как правило, занимает значительно больше времени, чем при митозе. Кроме того, клетки могут пребывать в стадии мейотической профазы I несколько дней, месяцев и даже лет, в зависимости от вида организма и типа образующихся гамет. (Эта растянутая фаза первого деления мейоза традиционно называется профазой, хотя она обладает большим сходством с фазой G2 обычного митоза в том отношении, что ядерная оболочка остается все время интактной и исчезает лишь тогда, когда начинается формирование нитей веретена, т.е. когда профаза I переходит в метафазу I.)
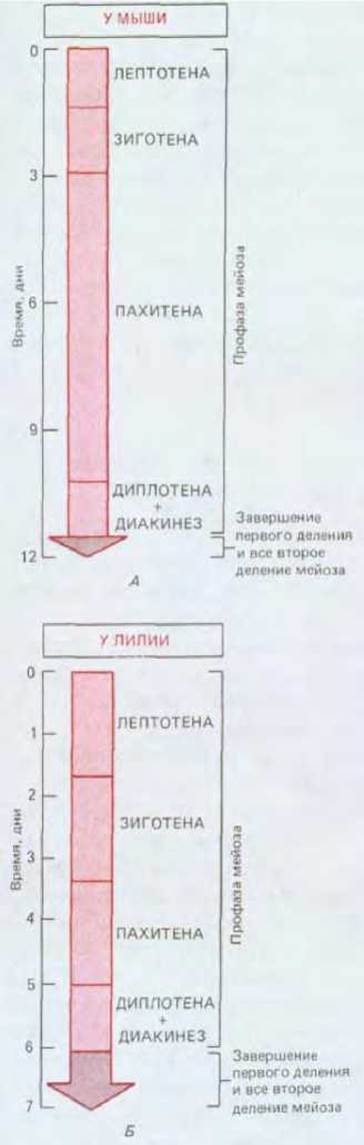
Рис. 15-20. Сравнение длительностей различных стадий мейоза, представленных на рис. 15-18. Показаны примерные временные интервалы для самца млекопитающего (мыши) и растения (лилии). Эти интервалы различны для женских и мужских гамет (яиц и спермиев) одного и того же вида и для одинаковых гамет разных видов. Например, мейоз у мужчины длится 24 дня, а у самца мыши 12 дней. Однако во всех случаях профаза первого деления мейоза намного продолжительнее, чем все остальные стадии, вместе взятые.
После окончания первого деления мейоза у двух дочерних ядер вновь образуются оболочки и начинается короткая интерфаза. В это время хромосомы несколько деспирализуются, однако, вскоре они опять конденсируются и начинается профаза II. Поскольку в этот период синтеза ДНК не происходит, создается впечатление, что у некоторых организмов хромосомы переходят непосредственно от одного деления к другому. Профаза II у всех организмов короткая: ядерная оболочка разрушается, когда формируется новое веретено, после чего, быстро сменяя друг друга, следуют метафаза II, анафаза II и телофаза II. Так же как и при митозе, у сестринских хроматид образуются кинетохорные нити, отходящие от центромеры в противоположных направлениях. В метафазной пластинке две сестринские хроматиды удерживаются вместе до анафазы, когда они разделяются благодаря внезапному расхождению их кинетохоров (рис. 15-19). Таким образом, второе деление мейоза сходно с обычным митозом (этого нельзя сказать о первом делении мейоза). Единственное существенное различие состоит в том, что здесь имеется по одной копии каждой хромосомы, а не по две, как в митозе.
Мейоз заканчивается формированием ядерных оболочек вокруг четырех гаплоидных ядер, образовавшихся в телофазе II (см. рис. 15-18, Б). Как мы увидим, у позвоночных к концу мейоза яйцеклетка уже полностью сформирована (а в некоторых случаях и оплодотворена), тогда как спермий еще только начинает свое развитие.
Заключение
При мейозе в результате двух последовательных клеточных делений, следующих за одним циклом репликации ДНК, из одной диплоидной клетки образуются четыре гаплоидные. У животных начальные фазы формирования яйцеклетки и сперматозоида сходны. В обоих случаях в мейозе доминирует профаза I, которая может занимать 90% всего времени мейоза. В этот период каждая хромосома состоит из двух тесно сближенных сестринских хроматид. Кроссинговер (перекрест) между хромосомами осуществляется на стадии пахитены в профазе I, когда конъюгация каждой пары гомологичных хромосом закрепляется синаптонемальным комплексом. Как полагают, каждый перекрест происходит при участии крупного рекомбинационного узелка и приводит к образованию хиазмы, сохраняющейся вплоть до анафазы I. В результате первого деления мейоза в каждую дочернюю клетку попадает по одной хромосоме из каждой пары гомологов, состоящих в это время из соединенных сестринских хроматид. Затем без репликации ДНК быстро протекает второе деление, при котором каждая сестринская хроматида попадает в отдельную гаплоидную клетку.