Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Половые клетки и оплодотворение
Гаметы
У эмбрионов всех позвоночных на ранней стадии развития определенные клетки обособляются как предшественники будущих гамет. Такие первичные половые клетки мигрируют в развивающиеся гонады (яичники у самок, семенники у самцов), где после периода митотического размножения претерпевают мейоз и дифференцируются в зрелые гаметы. Затем слияние яйцеклетки и спермия после спаривания инициирует процесс развития эмбриона, у которого, в свою очередь, формируются первичные половые клетки, т. е. открывается новый цикл.
Пока не ясно, по какой именно причине определенные клетки у зародыша млекопитающего превращаются в половые клетки, но известно, что, по крайней мере, у одного организма определяющим фактором служит какой-то компонент (или компоненты) цитоплазмы яйца: у дрозофилы специфическая область цитоплазмы - полярная плазма, расположенная на заднем полюсе яйца, - содержит мелкие гранулы, богатые РНК (полярные гранулы); клетки, образующиеся в этой части яйца и содержащие полярные гранулы, становятся первичными половыми клетками и, в конечном счете, мигрируют в гонады, где из них развиваются гаметы. Если полярную плазму ввести в передний полюс яйца, то клетки, которые должны были стать соматическими, превратятся в половые.
15.3.1. У высших животных яйцеклетка - это единственная клетка, из которой может развиться новая особь
По крайней мере, в одном отношении яйцеклетки - самые удивительные из всех животных клеток: будучи активированы, они могут дать начало целому новому организму, причем иногда для этого достаточно нескольких дней или недель. У высших животных это исключительная привилегия яйцеклеток. Активация обычно является следствием слияния яйцеклетки со спермием при оплодотворении, хотя, как мы позже увидим, во многих случаях яйцеклетку можно активировать иными, зачастую поразительно простыми способами (см. разд. 15.4.3). Активация яйцеклетки запускает программу развития, постепенное развертывание которой приводит к образованию новой особи.
У большинства животных, не относящихся к млекопитающим, ранний этап развития яйцеклетки сводится главным образом к ряду быстрых клеточных делений, или дроблению, при котором общая масса эмбриона остается, как правило, неизменной. В процессе дробления яйца образующиеся клетки постепенно становятся все меньше, пока не достигнут обычной величины зрелой соматической клетки. Хотя на ранних стадиях дробления синтезируются огромные количества ДНК и белков, в это время нет необходимости в синтезе РНК (в транскрипции генов). Это объясняется тем, что еще до оплодотворения в клетках накапливаются огромные резервы информационных РНК, рибосом, транспортных РНК и всех предшественников, необходимых для синтеза макромолекул, а также особых информационных РНК, кодирующих белки, которые необходимы для протекания ранних стадий развития.
Особенно большие запасы питательных веществ требуются тем яйцеклеткам, которые проходят длительный период эмбрионального развития вне родительского организма; в связи с этим яйца земноводных и птиц значительно крупнее, чем, например, яйцеклетки млекопитающих. Пресноводные и морские беспозвоночные, развивающиеся из небольших яйцеклеток вне родительского организма (такие, как морские ежи), обычно быстро превращаются в способных к самостоятельному питанию личинок.
Во взрослом организме из яйцеклетки может образоваться клетка любого типа. Тем не менее, ее отнюдь нельзя считать недифференцированной клеткой. Она в высшей степени специализирована для выполнения одной-единственной функции - построения новой особи. Прежде чем обсуждать вопрос о том, как протекает развитие яйцеклетки до того момента, когда она становится готова к оплодотворению, рассмотрим вкратце некоторые из ее уникальных особенностей.
15.3.2. Яйцеклетки представляют собой высокоспециализированные клетки, способные к независимому развитию и обладающие большими запасами питательных веществ и совершенной оболочкой [10]
Наиболее очевидная отличительная черта яйцеклетки - это ее большие размеры. Типичная яйцеклетка имеет сферическую или овальную форму, а диаметр ее составляет у человека и морского ежа около 100 мкм, у лягушек и рыб от 1 до 2 мм, а у птиц и рептилий измеряется сантиметрами (напомним, что величина типичной соматической клетки всего лишь около 20 мкм) (рис. 15-21 и 15-22). Столь же внушительными могут быть размеры ядра; например, в яйце лягушки величиной 1500 мкм диаметр ядра составляет около 400 мкм; в преддверии быстрых делений, следующих сразу за оплодотворением, в таком ядре откладываются запасы белков.
Потребность клетки в питательных веществах удовлетворяет в основном желток - материал протоплазмы яйца, богатый липидами и белками. Он обычно содержится в дискретных образованиях, называемых желточными гранулами. В яйцеклетках, развитие которых протекает вне материнского организма и приводит к формированию крупных животных, желток может занимать более 95% всего объема, тогда как у млекопитающих, чьи эмбрионы получают большую часть питательных веществ от матери, объем желтка составляет менее 5% объема яйцеклетки.
Другой важной специфической структурой яйцеклетки является наружная яйцевая оболочка покров из особого неклеточного вещества, состоящего в основном из гликопротеиновых молекул, часть которых секретирует сама яйцеклетка, а другую часть - окружающие клетки. У многих видов оболочка имеет внутренний слой, непосредственно прилегающий к плазматической мембране яйцеклетки и называемый у млекопитающих zona pellucida (рис. 15-23), а у других позвоночных и беспозвоночных (например, у морского ежа) - вителлиновым слоем. Этот слой защищает яйцеклетку от механических повреждений; в некоторых яйцеклетках он действует также как видоспецифический барьер для спермиев, позволяющий проникать внутрь только спермиям того же вида или очень близких видов (см. разд. 15.4.2). У животных, не относящихся к млекопитающим, соседние клетки часто выделяют добавочные оболочки, покрывающие вителлиновый слой. Например, когда яйца лягушки проходят из яичника по яйцеводу (трубка, по которой яйца выводятся наружу), их оболочка приобретает несколько дополнительных слоев из студнеобразного вещества, выделяемого эпителиальными клетками яйцевода. Аналогичным образом у куриного яйца при прохождении его по яйцеводу (после оплодотворения) появляются «белок» и твердая скорлупа, тогда как яйца насекомых покрываются тонкой прочной оболочкой, получившей название хориона и выделяемой фолликулярными клетками, окружающими каждое яйцо.
Многие яйцеклетки (в том числе и яйцеклетки млекопитающих) содержат специализированные секреторные пузырьки, находящиеся под плазматической мембраной в наружном, или кортикальном, слое цитоплазмы (рис. 15-24). При активации яйцеклетки спермием эти кортикальные гранулы высвобождают содержимое путем экзоцитоза, в результате свойства яйцевой оболочки изменяются таким образом, что через нее уже не могут проникнуть внутрь яйцеклетки другие спермин (см. разд. 15.4.5).
В то время как кортикальные гранулы обычно равномерно распределены во всем кортексе яйцеклетки, другие компоненты цитоплазмы могут располагаться крайне асимметрично. Например, в яйце лягушки большая часть желтка находится на одном (вегетативном) полюсе, тогда как ядро располагается ближе к противоположному (анимальному) полюсу. Полярность яйцеклетки, которая зависит главным образом от внешних условий, часто определяет полярность развивающегося эмбриона (см. разд. 16.1.1).

Рис. 15-21. Три различных яйца в натуральную величину. Диаметр яйцеклетки человека составляет 0,1 мм.
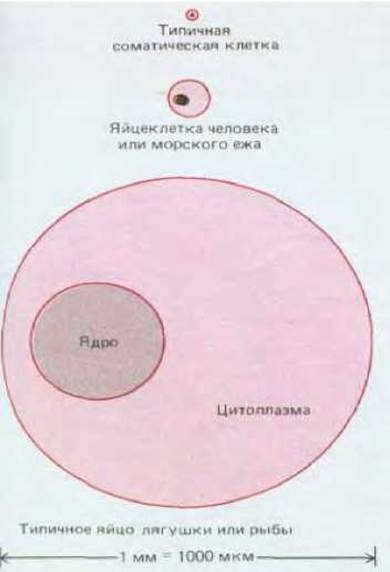
Рис. 15-22. Относительные размеры различных яйцеклеток по сравнению с величиной типичной соматической клетки.
15.3.3. Яйцеклетка проходит в своем развитии несколько стадий [10, 11]
Развивающаяся яйцеклетка называется ооцитом; ее дифференцировка в зрелую яйцеклетку (или яйцо), включающая в себя ряд высокоспециализированных стадий, отличается от обычных клеточных циклов. Мы уже знаем, что вместо того, чтобы продолжать делиться посредством обычного митоза, половые клетки осуществляют два последних деления путем более сложного процесса-мейоза. Более того, у ооцитов выработались специальные механизмы, позволяющие им приостанавливать ход мейоза: ооциты на длительный период задерживаются в профазе 1, увеличиваясь в это время в размерах, а позже они в преддверии оплодотворения во многих случаях временно останавливают свое развитие на стадии метафазы II.
Хотя детали развития яйцеклетки (оогенеза) различаются у разных видов, основные стадии сходны (рис. 15-25). Первичные половые клетки мигрируют в формирующуюся гонаду и превращаются в оогонии; после периода митотического размножения оогонии дифференцируются в ооциты первого порядка, которые приступают к первому делению мейоза. Происходит репликация ДНК, и каждая хромосома состоит после этого из двух хроматид; гомологичные хромосомы конъюгируют по всей своей длине, и между хроматидами спаренных хромосом осуществляется кроссинговер. На этой стадии профаза приостанавливается на то или иное время, которое может составлять от нескольких дней до многих лет в зависимости от вида организма. Во время этой затянутой профазы (или, в некоторых случаях, при наступлении половой зрелости) ооциты первого порядка приобретают наружные оболочки и кортикальные гранулы, накапливают рибосомы, желток, гликоген, липиды и информационную РНК, которая впоследствии направляет синтез белков, необходимых для ранних стадий роста эмбриона и подготовки к развертыванию программы развития. Во многих ооцитах такая активность отражается на видимой структуре все еще спаренных хромосом: они деспирализуются и образуют боковые петли, приобретая характерный вид «ламповых щеток», свойственный хромосомам, активно осуществляющим синтез РНК (см. разд. 9.2.4).
Следующая фаза развития, называемая созреванием яйцеклетки, начинается лишь с наступлением половой зрелости. Под влиянием гормонов (см. ниже) происходит первое деление мейоза: хромосомы снова конденсируются, ядерная оболочка исчезает (этот момент обыкновенно принимают за начало созревания), и в анафазе I реплицированные гомологичные хромосомы расходятся в дочерние ядра, каждое из которых содержит теперь половину исходного числа хромосом. Но цитоплазма делится очень несимметрично, так что к концу первого деления получаются две клетки, резко различающиеся по величине: одна представлена маленьким полярным тельцем, а другая - большим ооцитом второго порядка, в котором заложены все возможности для развития. На этой стадии каждая из хромосом все еще состоит из двух сестринских хроматид; в таком виде хромосомы остаются вплоть до второго деления мейоза, когда хроматиды отделяются друг от друга в результате процесса, сходного с анафазой митоза (см. разд. 15.2.7). После окончательного расхождения хромосом в анафазе II цитоплазма большого ооцита второго порядка вновь делится асимметрично, что ведет к образованию зрелой яйцеклетки (или яйца) и еще одного маленького полярного тельца; при этом обе клетки получают гаплоидное число одиночных хромосом (см. рис. 15-25). Благодаря двум несимметричным делениям цитоплазмы ооциты сохраняют большую величину, хотя они и претерпели два деления мейоза. Все полярные тельца очень малы, и они постепенно дегенерируют.

Рис. 15-23. А. Микрофотографии яйцеклетки хомячка, полученные с помощью сканирующего электронного микроскопа. Хорошо видна zona pellucida; на фото Б эта оболочка (к которой прикрепилось много спермиев) частично отогнута и можно видеть лежащую под ней плазматическую мембрану яйцеклетки с многочисленными микроворсинками. (М. Pillips, J. Ultrastruct. Res., 72, 1-12, 1980.)
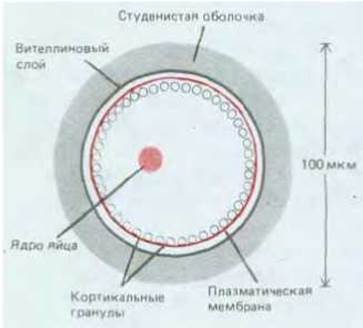
Рис. 15-24. Схематический разрез яйца морского ежа; видно расположение гранул. Обратите внимание на то, что вителлиновый слой покрыт студенистой оболочкой, толщина которой доходит до 30 мкм.
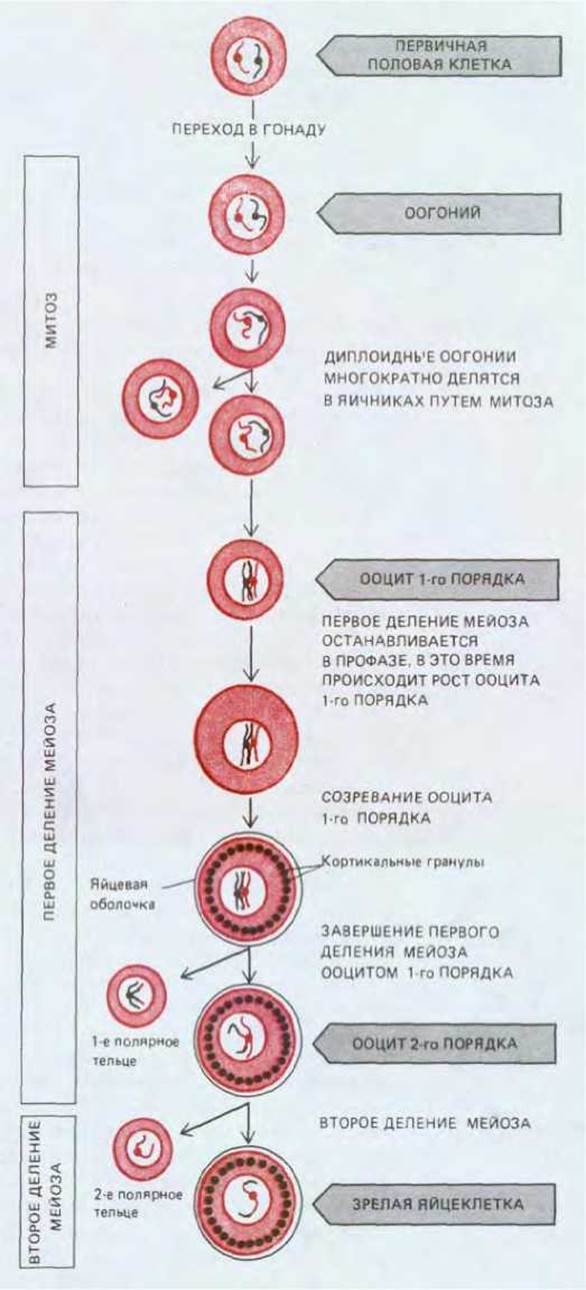
Рис. 15-25. Различные стадии оогенеза. Из первичных половых клеток, мигрирующих в яичник на ранней стадии эмбриогенеза, развиваются оогонии. После ряда митотических делений оогонии приступают к первому делению мейоза, и на этой стадии их называют уже ооцитами первого порядка. У млекопитающих ооциты первого порядка формируются очень рано и остаются на стадии профазы I (у человека это происходит между 3м и 8-м месяцами эмбрионального развития) до тех пор, пока самка не достигнет половой зрелости. После этого под влиянием гормонов периодически созревает небольшое число ооцитов, которые завершают первое деление мейоза и превращаются в ооциты второго порядка; последние претерпевают второе деление мейоза и становятся зрелыми яйцеклетками. Стадия, на которой яйцеклетка выходит из яичника и оплодотворяется, у разных животных различна. У большинства позвоночных созревание ооцитов приостанавливается на стадии метафазы II и ооцит второго порядка завершает мейоз лишь после оплодотворения. Все полярные тельца в конечном счете дегенерируют. Однако у большинства животных, в том числе и у млекопитающих, полярные тельца остаются внутри яйцевой оболочки, а у некоторых видов первое полярное тельце успевает до дегенерации претерпеть одно деление.
У большинства позвоночных созревание ооцитов продолжается до метафазы II и приостанавливается на этой стадии. При овуляции ооцит второго порядка освобождается из яичника и в том случае, когда происходит оплодотворение, завершает мейоз.
15.3.4. Яйцеклетка достигает крупных размеров благодаря специальным механизмам [10, 11]
Небольшой соматической клетке диаметром 10-20 мкм обычно требуется около суток, для того чтобы удвоить массу при подготовке к делению. Той же клетке при таких же скоростях синтеза макромолекул понадобилось бы очень много времени, чтобы достичь в тысячу раз большей массы, характерной для яйцеклетки млекопитающего (диаметр 100 мкм), или в миллион раз большей массы яйца насекомого (диаметр 1000 мкм). Между тем некоторые насекомые живут всего лишь несколько дней и ухитряются производить яйца, диаметр которых даже больше 1000 мкм. Ясно, что их яйцеклетки должны обладать особыми механизмами для достижения столь крупных размеров.
Одним из факторов, способствующих такому росту, является наличие в клетке дополнительных копий генов. У яйцеклеток многих животных завершение мейоза откладывается почти до самого конца созревания, так что эти яйцеклетки содержат удвоенный диплоидный набор хромосом в течение большей части периода их роста. Таким образом, они содержат вдвое больше ДНК для синтеза РНК, чем имеет средняя соматическая клетка в фазе клеточного цикла. В некоторых яйцеклетках процесс накопления дополнительной ДНК идет еще дальше, приводя к образованию большого числа добавочных копий определенных генов. В гл. 9 мы уже видели, что для образования достаточного числа рибосом, на которых происходит синтез белков, соматическим клеткам большинства организмов требуется от 10 до 500 копий генов рибосомной РНК. Поскольку яйцеклетки нуждаются в еще большем количестве рибосом для белкового синтеза на ранних стадиях эмбриогенеза, в яйцеклетках многих животных гены рРНК амплифицируются; так, в яйцах рыб и амфибий содержится 1-2 млн. копий таких генов (рис. 15-26).
Рост многих яиц в какой-то степени зависит от биосинтетической активности других клеток. Например, один из важных компонентов крупной яйцеклетки - желток - обычно синтезируется вне яичника и затем переносится в ооцит. У птиц, амфибий и насекомых белковые вещества желтка образуются в клетках печени (или их функциональных аналогах), которые выделяют эти вещества в кровь. Ооциты, находящиеся в яичниках, извлекают эти будущие белковые компоненты желтка из внеклеточной жидкости путем эндоцитоза при участии специфических рецепторов (см. рис. 6-72). Питанию ооцитов могут также способствовать вспомогательные клетки, находящиеся в яичнике. Эту функцию в оогенезе в зависимости от вида организма выполняют вспомогательные клетки яичника двух различных типов. У некоторых беспозвоночных имеются клетки-кормилицы: они обычно окружают яйцеклетку и соединены с ней цитоплазматическими мостиками, по которым макромолекулы могут прямо переходить в ее цитоплазму. Клетки-кормилицы синтезируют для яйцеклеток насекомых такие компоненты (рибосомы, мРНК, белки и т. п.), которые у позвоночных яйцеклетка производила бы для себя сама.

Рис. 15-26. Микрофотография изолированного ядра из яйца лягушки. Ядро окрашено крезиловым фиолетовым, чтобы можно было видеть множество ядрышек результат многократной амплификации генов рибосомной РНК. (D. D. Brown., I. Dawid, Science, 160, 273-275, 1968.)

Рис. 15-27. Эта схема показывает, как из одного оогония Drosophila образуется 15 клеток-кормилиц и один ооцит; все они связаны между собой цитоплазматическими мостиками. При каждом митозе все клетки однократно делятся: при первом митозе из клетки 1 образуются клетки 1 и 2; при втором митозе из клетки 1 образуются клетки 1 и 3, а из клетки 2-клетки 2 и 4 и т.д. Поскольку цитоплазматические мостики образуются во всех тех местах, где остатки веретена деления связывали во время телофазы две дочерние клетки, эти мостики соединяют лишь те клетки, которые образовались в результате общего митоза. В яйцеклетку превращается только клетка 1 или 2; возможно, это связано с тем, что только эти клетки соединены межклеточными мостиками с четырьмя другими. Необычной особенностью таких делений является то, что размер клетки не удваивается перед митозом, так что с каждым делением клетка становится все меньше и меньше. Позже, во время созревания яйца, клетки-кормилицы становятся чрезвычайно крупными; они образуют большие количества макромолекул и таких органелл, как рибосомы и митохондрии, и «накачивают» их внутрь ооцита по цитоплазматическим мостикам.
У некоторых видов клетки-кормилицы происходят из того же оогония, из которого образуется соединенный с ними ооцит. Например, у дрозофилы оогоний претерпевает четыре митотических деления, в результате которых образуется 16 клеток. Одна из этих клеток становится яйцом, а другие превращаются в клетки-кормилицы и остаются соединенными друг с другом и с яйцом цитоплазматическими мостиками (рис. 15-27). В клетках-кормилицах происходит многократная репликация ДНК без деления самой клетки, поэтому каждая клетка постепенно достигает очень больших размеров, а количество ДНК в ней в тысячу раз превосходит обычную величину (такая ДНК находится в политенных хромосомах; см. разд. 9.2.5). Все 15 клеток-кормилиц, содержащих сотни и тысячи эквивалентов генома, синтезируют вещества, необходимые для одной-единственной яйцеклетки.
Еще один вид клеток, которые помогают обеспечить питание развивающихся ооцитов,- это фолликулярные клетки, имеющиеся как у позвоночных, так и у беспозвоночных. Они расположены вокруг ооцита в виде эпителиального слоя (рис. 15-28) и связаны с ним щелевыми контактами, через которые могут проходить малые молекулы, но не макромолекулы (разд. 14.1.5). Хотя подобные клетки не могли бы через эти контакты снабжать ооцит готовыми макромолекулами, возможно, что они помогают обеспечить его малыми молекулами-предшественниками, из которых образуются макромолекулы. Кроме того, фолликулярные клетки часто выделяют макромолекулы, которые либо входят в состав оболочки яйцеклетки, либо путем эндоцитоза поступают в растущий ооцит.
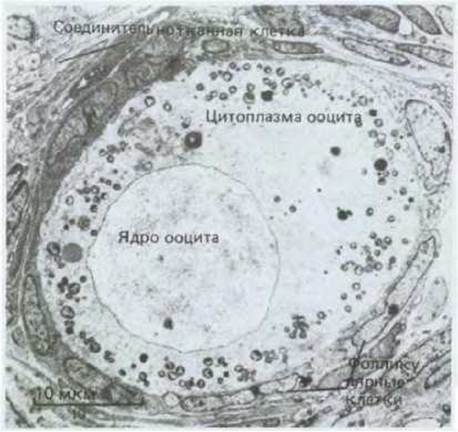
Рис. 15-28. Примордиальный фолликул кролика, состоящий из центрального ооцита, который окружен одним слоем уплощенных фолликулярных клеток (электронная микрофотография). Примордиальный фолликул заключен в базальную мембрану, плохо различимую на фотографии, которая в свою очередь окружена соединительной тканью яичника. Обратите внимание, что на этой стадии развития ооцит первого порядка не имеет zona pellucida и кортикальных гранул. (J. Van Blerkom, P. Motta, Cellular Basis of Mammalian Reproduction, Baltimore: Urban and Schwarzenberg, 1979.)
Благодаря вспомогательным клеткам, окружающим ооцит, становится возможным продолжение его развития; и у позвоночных, и у беспозвоночных именно вспомогательные клетки реагируют на действие полипептидных гормонов (гонадотропинов), в результате чего прерывается задержка ооцитов в профазе I и они становятся готовыми к дальнейшему созреванию.
15.3.5. Созревание ооцитов у позвоночных инициируется уменьшением внутриклеточной концентрации циклического AMP [12]
У многих животных завершение ооцитом мейоза I происходит лишь с наступлением половой зрелости, когда в крови появляются гонадотропины. Эти полипептидные гормоны стимулируют вспомогательные клетки яичника, побуждая их к выделению вторичного медиатора, который в свою очередь воздействует на ооциты и индуцирует процесс их созревания. Для большинства позвоночных созреванием яйцеклетки считается процесс развития ооцита первого порядка, охватывающий период времени от первого деления мейоза до новой задержки ооцита в метафазе II; на этой стадии теперь уже ооцит второго порядка остается до оплодотворения, под влиянием которого происходит завершение мейоза (рис. 15-25).
Процесс созревания яйцеклетки лучше всего изучен у амфибий. У этих животных гонадотропины, находящиеся под контролем гипофиза, действуя на окружающие ооциты фолликулярные клетки, инициируют выделение последними стероидного гормона прогестерона. Подобно другим стероидным гормонам, прогестерон диффундирует через плазматические мембраны большинства клеток-мишеней и связывается с внутриклеточными рецепторными белками, регулирующими транскрипцию специфических генов (см. разд. 12.2.1). Однако в созревании ооцита прогестерон, по-видимому, участвует иначе: полагают, что он связывается с рецепторными белками плазматической мембраны. При этом происходит инактивация аденилатциклазы плазматической мембраны ооцита, в результате чего снижается концентрация циклического AMP в цитозоле и соответственно активность сАМР-зависимой протеинкиназы (А-киназы, см. разд. 12.4.1).
Имеются убедительные свидетельства того, что снижение активности А-киназы является необходимым и достаточным условием для запуска программы созревания ооцита. Микроинъекции каталитически активного фрагмента киназы, активного в отсутствие циклического AMP (см. разд. 12.4.2), ингибируют созревание, индуцируемое прогестероном, тогда как микроинъекции специфического ингибитора киназы инициируют созревание даже при отсутствии прогестерона. Эксперименты на мышиных ооцитах, проводившиеся также с помощью метода микроинъекций, показали, что у млекопитающих действует аналогичный внутриклеточный механизм передачи сигналов. Субстраты А-киназы пока не выявлены, однако полагают, что ими могут быть фосфопротеины, входящие в систему прерывания мейотического деления I, ответственную за задержку ооцита в профазе I. По имеющейся гипотезе, активность таких фосфопротеинов определяется фосфорилированием, осуществляемым А-киназой; падение уровня циклического AMP снимает блокировку мейоза и инициирует его продолжение.
15.3.6. Созревание ооцитов связано с активацией фактора инициации М-фазы [13]
Пока еще не выяснено, как развиваются события после падения уровня циклического AMP, но в конце концов они приводят к активации белкового комплекса, называемого фактором инициации М-фазы (ФИМ); полагают, что он необходим для выхода из профазы I мейоза. В гл. 13 говорилось о том, что ФИМ крайне мало изменился в процессе эволюции эукариот. Его ключевая роль в обычном цикле клеточного деления обусловлена тем, что он инициирует переход клетки из G2 в М-фазу (см. разд. 13.1.10). Как уже упоминалось, профаза I мейоза, несмотря на свое традиционное название, очень похожа на фазу G2 обычного клеточного цикла: ДНК уже реплицировалась и является активной в отношении транскрипции, ядерная оболочка интактна, а митотическое веретено деления еще не сформировалось. Более того, подобно переходу из G2 в М-фазу обычной делящейся клетки, переход от профазы I к М-фазе мейоза запускается ФИМ. В действительности ФИМ был впервые обнаружен в ооцитах лягушки в качестве фактора, инициирующего созревание. Зрелые ооциты лягушки задерживаются на стадии метафазы II, когда уровень ФИМ высок (см. далее). Если небольшую часть цитоплазмы такого зрелого ооцита ввести в незрелый ооцит, то под воздействием содержащегося в инъекции ФИМ нарушится целостность мембраны и начнется конденсация хромосом, т. е. будут наблюдаться эффекты, характерные для М-фазы и свидетельствующие о зрелости ооцита.
Замечательное свойство ФИМ состоит в том, что его можно неограниченно долго последовательно переносить из одного ооцита в другой без потери активности, несмотря на то что при подобных последовательных переносах разбавление раствора с ФИМ достигает огромных значений. Это означает, что инъекции ФИМ стимулируют синтез в реципиенте-ооците еще больших количеств ФИМ. Амплификация происходит даже в безъядерных ооцитах и в присутствии ингибиторов синтеза белков; такой факт указывает на то, что ФИМ скорее всего активирует уже существующие ФИМ, а не индуцирует синтез новых ФИМ.
Какой механизм лежит в основе такой автокаталитической реакции саморазмножения? Несмотря на то что к настоящему времени ФИМ еще не изучен досконально, имеются свидетельства того, что он фосфорилирован, что фосфорилированию принадлежит важная роль в возбуждении активности ФИМ и что по меньшей мере один из компонентов ФИМ является протеинкиназой. Таким образом, ФИМ может сам себя фосфорилировать и, следовательно, активировать. В ооцитах лягушки инактивация А-киназы под действием прогестерона инициирует сложную последовательность событий (сопряженную с синтезом белков), которая в конце концов приводит к активации небольших количеств ФИМ, которые, в свою очередь, активируют новые ФИМ. Интенсивное фосфорилирование структурных белков ядерных оболочек и хромосом характерно для процессов, наблюдаемых в М-фазе (см. разд. 13.5.11), поэтому напрашивается предположение о том, что ФИМ синтезирует еще и неизвестную протеинкиназу, которая вызывает такого рода дальнейшие изменения (рис. 15-29).
Считают, что циклические изменения активности ФИМ, по-видимому, являются решающим фактором регуляции обычного митотического клеточного цикла, определяя моменты вступления клетки в митоз и выхода из него (см. разд. 13.1.10). По-видимому, при созревании ооцитов ФИМ претерпевает аналогичные циклические изменения. Увеличение количества ФИМ сначала запускает прохождение ооцита через профазу и вступление его в метафазу I деления мейоза. Затем последующая инактивация ФИМ позволяет ооцитам завершить деление ядра, а новая активация ФИМ инициирует переход клетки в М-фазу II деления мейоза. Полагают, что система задержки II мейотического деления (ее иногда отождествляют с цитостатическим фактором, или ЦСФ, свойства которого слабо изучены) на этой стадии предотвращает инактивацию ФИМ, приостанавливая таким образом зрелые яйцеклетки на стадии метафазы П. Оплодотворение, возможно, благодаря повышению концентрации Са2+ в цитозоле (см. разд. 15.4.5), позволяет ооцитам завершить II деление мейоза и вместе с последовательными сериями активации и инактивации ФИМ, определяющими прохождение клеток через повторяющиеся митотические деления, запускает процесс развития эмбриона.

Рис. 15-29. Модель, демонстрирующая, каким образом вслед за воздействием прогестерона фактор инициации М-фазы (ФИМ) может индуцировать переход яйца лягушки из профазы 1 в метафазу. По предложенной гипотезе, прогестерон косвенно вызывает образование соответствующего фермента, который активирует небольшие количества ФИМ. Это приводит к активации не только больших количеств ФИМ (многократно усиленный отклик), но и протеинкиназ, фосфорилирующих ядерные мембраны и хромосомные белки. В результате целостность ядерной оболочки нарушается, хромосомы конденсируются и, таким образом, клетка вступает в метафазу. Последующая инактивация ФИМ приводит к тому, что воссоздается ядерная оболочка, хромосомы деконденсируются и, таким образом, клетка может вступить во второе деление мейоза (на рисунке не показано). Возможно, аналогичный механизм лежит в основе процесса созревания ооцитов млекопитающих. Один из компонентов ФИМ идентифицирован: это протеинкиназа. Она гомологична протеинкиназе дрожжей, кодируемой геном cdc2/28 и играющей ключевую роль в регуляции клеточного цикла дрожжевых клеток.
15.3.7. Большая часть ооцитов, не созревая, погибает в яичнике человека [14]
Несмотря на то что молекулярные механизмы, лежащие в основе важнейших моментов в процессе развития и созревания ооцитов, по всей видимости, весьма сходны у всех позвоночных, временные параметры этих процессов и характер запускающих их внеклеточных сигналов могут варьировать весьма значительно. Например, у человека эти сигналы значительно более сложны и гораздо менее изучены, чем у амфибий. Более того, количества зрелых яйцеклеток, образующихся у представителей этих двух видов, различаются поразительным образом. У самки лягушки в каждый сезон размножения на протяжении всего репродуктивного периода жизни из оогоний образуются ооциты; сотни их делятся и претерпевают овуляцию под действием гормонов. У женщины же лишь в первые месяцы внутриутробного развития из оогоний образуется несколько миллионов ооцитов, и все они, за исключением нескольких сотен, не созревая, погибают в яичнике.
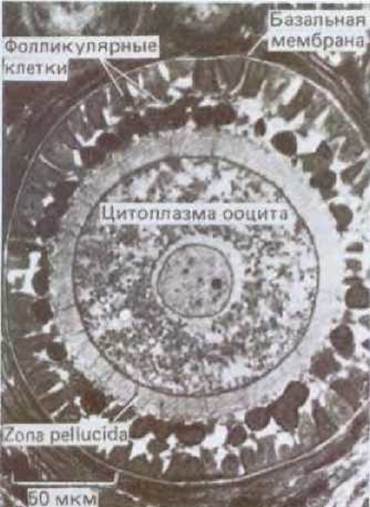
Рис. 15-30. Микрофотография созревающего фолликула в яичнике крольчихи. Ооцит первого порядка крупнее, чем более молодые ооциты, представленные на рис. 15-28, и уже имеет кортикальные гранулы (на этом фото они плохо видны), а также узкую zona pellucida; ооцит окружен слоями фолликулярных клеток. Внутренние фолликулярные клетки имеют отростки, которые проходят сквозь zona pellucida и образуют щелевые контакты с ооцитом (правда, на фотографии все эти структуры видны нечетко). Весь фолликул окружен базальной мембраной. (J. Van Blerkom, P. Motta. Cellular Basis of Mammalian Reproduction, Baltimore, Urban and Schwarzenberg, 1979.)
Ооциты первого порядка у новорожденной девочки останавливаются в профазе I мейоза (подобно ооцитам первого порядка у амфибий), и большая их часть окружена одним слоем фолликулярных клеток; такой ооцит вместе с этими фолликулярными клетками представляет собой примордиальный фолликул (рис. 15-28). За некоторое время до рождения небольшая доля примордиальных фолликулов периодически начинает расти, превращаясь в развивающиеся фолликулы: их клетки увеличиваются и размножаются, образуя вокруг ооцита первого порядка многослойную оболочку; сам ооцит, оставаясь в профазе 1, растет, и у него формируются zona pellucida и кортикальные гранулы (рис. 15-30). В некоторых из развивающихся фолликулов образуется наполненная жидкостью полость, или antrum, при этом они превращаются в антральные фолликулы (рис. 15-31). Чем индуцируется первоначальный рост отдельных примордиальных фолликулов и превращение их в развивающиеся фолликулы, пока неясно. Однако продолжение развития, вероятно, зависит от гонадотропных гормонов гипофиза [в основном от фолликулостимулирующего гормона (ФСГ)] и от эстрогенов, секретируемых самими фолликулярными клетками.
Все развивающиеся фолликулы продолжают расти и в конце концов либо дегенерируют, либо в процессе овуляции высвобождают находящиеся внутри них ооциты. Однако созревание ооцита (прохождение через профазу I) и овуляция происходят лишь после наступления половой зрелости. С этого времени раз в месяц (примерно в середине менструального цикла) резкий подъем уровня другого гонадотропина - лютеинизирующего гормона (ЛГ), выделяемого гипофизом, - стимулирует рост в яичнике 15-20 тыс. развивающихся фолликулов. Из всех этих фолликулов лишь один завершает свое развитие: заключенный в нем ооцит первого порядка созревает, доходит до метафазы II деления мейоза и останавливается на этой стадии. Единственный стимулированный фолликул тем временем быстро увеличивается в размерах и разрывается на поверхности яичника, освобождая находящийся внутри него ооцит второго порядка, который к этому времени все еще остается заключенным в оболочку из фолликулярных клеток, внедренную в студ-необразный матрикс (рис. 15-32). Инициация завершения мейоза у освобожденного ооцита произойдет лишь в том случае, если он будет оплодотворен спермием не позже, чем через сутки (или чуть более того).
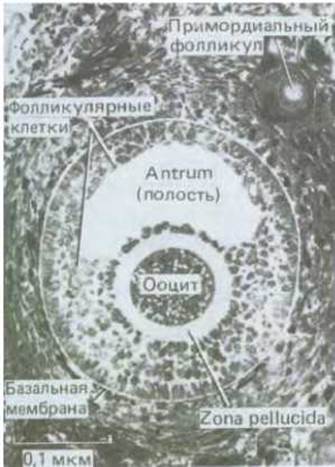
Рис. 15-31. Микрофотография антрального фолликула в яичнике крольчихи. Теперь здесь имеется много слоев фолликулярных клеток. Ядра ооцитов не попали в плоскость сечения и поэтому не видны на фотографии. Для сравнения обратите внимание на примордиальный фолликул значительно меньших размеров в верхнем правом углу. (J. Van Blerkom, P. Motta. Cellular Basis of Mammalian Reproduction, Baltimore, Urban and Schwarzenberg, 1979.)
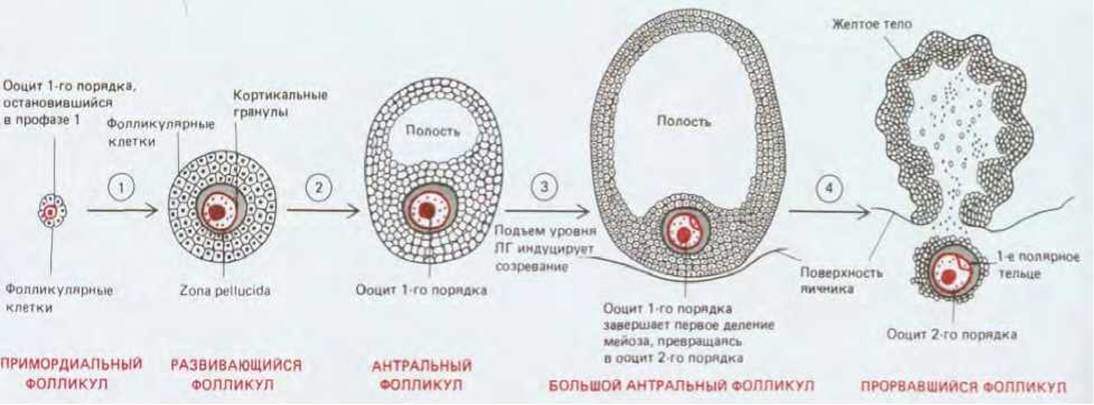
Рис. 15-32. Здесь схематически показаны стадии развития ооцита у человека. 1. До рождения небольшая доля примордиальных фолликулов последовательно начинает расти, и эти фолликулы называют теперь развивающимися. 2. Некоторые из развивающихся фолликулов накапливают жидкость, превращаясь в антральные фолликулы. 3. С наступлением половой зрелости раз в месяц волна лютеинизирующего гормона (ЛГ) побуждает примерно 20 антральных фолликулов к ускоренному росту, однако завершает созревание и претерпевает овуляцию лишь один из этих фолликулов; ооцит первого порядка, находящийся в этом фолликуле, завершает первое деление мейоза, образуя полярное тельце и превращаясь в ооцит второго порядка (рис. 15-25). 4. Ооцит второго порядка, который был остановлен на стадии метафазы мейоза II, вместе с полярным тельцем и частью окружающих полярных клеток освобождается в тот момент, когда фолликул разрывается на поверхности яичника. Ооцит второго порядка завершает второе деление мейоза только в том случае, если он будет оплодотворен. После овуляции опустевший фолликул превращается в эндокринное образование (желтое тело), которое секретирует прогестерон, тем самым готовя матку к приему оплодотворенного ооцита. Если оплодотворение не наступает, желтое тело претерпевает обратное развитие, а внутренняя слизистая оболочка матки отслаивается и выводится из организма во время менструации.
Каким образом подъем уровня ЛГ, происходящий в середине менструального цикла, инициирует созревание ооцита? По-видимому, фолликулярные клетки, окружающие ооцит, обычно оказывают ингибирующее воздействие на его созревание, поскольку ооциты спонтанно возобновляют ход мейоза в том случае, когда от них удаляют антральные фолликулы и лишают среду, в которой культивируется ооцит, стимулирующих гормонов. Подъем уровня ЛГ может стимулировать созревание яйцеклетки путем устранения ингибирующего воздействия фолликулярных клеток.
Одна из загадочных особенностей созревания яйцеклетки у человека состоит в том, что лишь малая часть из многочисленных антральных фолликулов, имеющихся в яичниках к моменту ежемесячного выброса ЛГ, начинает расти, и из этих растущих фолликулов только один завершает созревание и освобождает ооцит: остальные обречены на дегенерацию. Вероятно, тотчас после того, как созревание «избранного» фолликула достигло определенной критической точки, вступает в действие некий механизм обратной связи, благодаря которому никакой другой фолликул не может созреть и претерпеть овуляцию в том же цикле. Это означает, что в течение примерно 40-летнего репродуктивного периода у женщины выделяется лишь 400-500 яйцеклеток. Оставшиеся 3 млн. ооцитов первого порядка, имевшиеся в организме к моменту рождения, дегенерируют. До сих пор остается загадкой, почему огромное число яйцеклеток формируется лишь для того, чтобы погибнуть в яичниках.
Ооциты второго порядка, освобождающиеся из яичника (при овуляции) к концу репродуктивного периода, образуются из ооцитов первого порядка, которые провели в остановленной профазе I от 40 до 50 лет. Дефекты, возникающие за это время в яйцеклетке, могут быть причиной высокой частоты генетических аномалий среди детей, рожденных немолодыми женщинами. Например 1% детей у женщин старше 40 лет страдает синдромом Дауна, обусловленным трисомией по 21-й хромосоме (следствие нерасхождения соответствующих гомологов при делении ядра созревающего ооцита в мейозе I) (рис. 15-33).
15.3.8. Спермии отлично приспособлены для внесения своей ДНК в яйцеклетку [15]
У большинства видов имеется два вида гамет, поразительно отличающихся друг от друга. Если яйцеклетка - одна из самых крупных клеток организма, то спермий (или сперматозоид) обычно меньше всех других клеток. Яйцеклетка и сперматозоид наилучшим образом, но по-разному, приспособлены для распространения имеющихся в них генов. Яйцеклетка обеспечивает выживаемость материнских генов за счет больших запасов веществ, необходимых для роста и развития, а также благодаря прекрасной защитной оболочке. В противоположность этому сперматозоид обычно наилучшим образом справляется с задачей оплодотворения и сохранения отцовских генов благодаря тому, что является исключительно подвижным и обладает обтекаемой формой, позволяющей ему быстро двигаться. Сперматозоиды действуют в условиях жесточайшей конкуренции, и подавляющее их большинство погибает в процессе исполнения своей миссии: из миллиардов сперматозоидов, выделяемых мужчиной в течение репродуктивного периода жизни лишь считанные единицы достигают своей цели и оплодотворяют яйцеклетку.
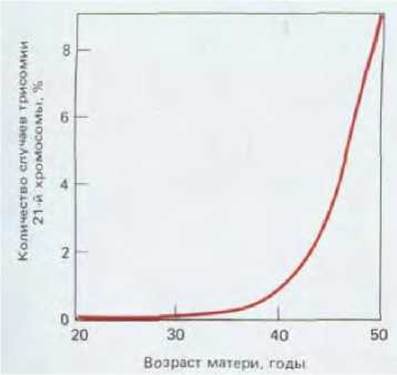
Рис. 15-33. Увеличение числа случаев трисомии по 21-й хромосоме (синдрома Дауна) у новорожденных в зависимости от возраста матери. (По: L. Mastroianni, Jr. and J. D. Digger, eds. Fertilization and Embryonic Development in Vitro, p. 260, Plenum, 1981.
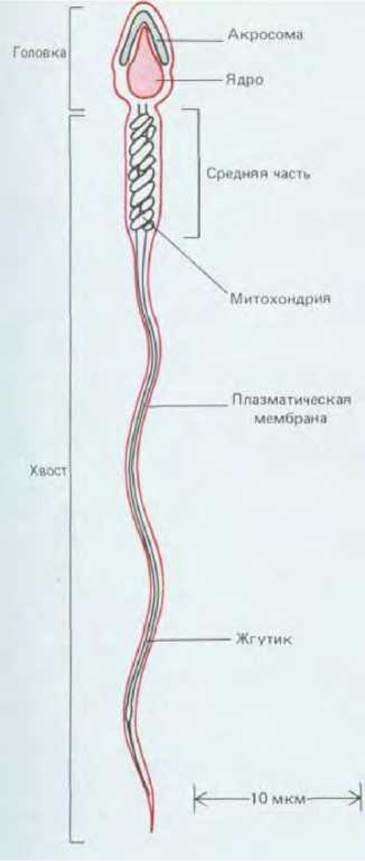
Рис. 15-34. Сперматозоид человека, схематически представленный в продольном разрезе.
Типичный спермий (рис. 15-34) - это клетка, снабженная мощным жгутиком, благодаря которому она движется в жидкой среде; при этом спермий «избавлен от всего лишнего» и не обременен такими органеллами, как рибосомы, эндоплазматический ретикулум или аппарат Гольджи, присутствия которых не требуется для передачи ДНК яйцеклетке. С другой стороны, спермии содержат много митохондрий, расположенных в тех местах, где они могут наиболее эффективно снабжать энергией жгутик. Спермий обычно состоит из двух морфологически и функционально различающихся частей, заключенных в единую плазматическую мембрану: из головки, содержащей необычайно сильно уплотненное гаплоидное ядро, и хвоста, который продвигает всю клетку по направлению к яйцу и способствует прохождению головки через яйцевую оболочку. ДНК в ядре неактивна и исключительно плотно упакована, так что объем ее доведен до минимума. Хромосомы многих сперматозоидов обходятся даже без гистонов, свойственных соматическим клеткам, вместо этого здесь имеются простые белки, обладающие большим положительным зарядом.
В головке спермия, вплотную к передней части ядерной мембраны, располагается специализированный секреторный пузырек, называемый акросомой (рис. 15-34). Этот пузырек содержит гидролитические ферменты, позволяющие спермию проникнуть сквозь наружные яйцевые оболочки. Когда его головка приходит в контакт с яйцеклеткой, содержимое акросомы высвобождается путем экзоцитоза (так называемая акросомальная реакция). У беспозвоночных в ходе этой реакции выводятся наружу специфические белки, прочно прикрепляющие спермий к яйцевой оболочке (см. разд. 15.4.2).
Подвижный хвост спермия представляет собой длинный жгутик, аксонема которого начинается от базального тельца, расположенного сразу за ядром. Ранее (разд. 11.3.5) уже говорилось, что аксонема состоит из двух одиночных центральных микротрубочек, окруженных девятью равноотстоящими друг от друга дублетами микротрубочек. У некоторых животных (в том числе у млекопитающих) жгутик спермия отличается от других жгутиков тем, что вокруг его аксонемы лежат еще девять внешних плотных волокон неизвестного состава, так что вместо обычной схемы 9 + 2 мы имеем здесь схему 9 + 9 + 2 (рис. 15-35 и 15-36). Эти плотные волокна жестки и не могут сокращаться; неизвестно, способствуют ли они энергичным изгибам жгутика, которые вызываются скольжением соседних дублетов микротрубочек относительно друг друга (разд. II.3.6). Энергию для движения жгутика поставляет гидролиз АТР, синтезируемого высокоспециализированными митохондриями, которые находятся именно там, где они больше всего нужны, т. е. в передней части хвоста (называемой средней частью сперматозоида) (рис. 15-34 и 15-35).
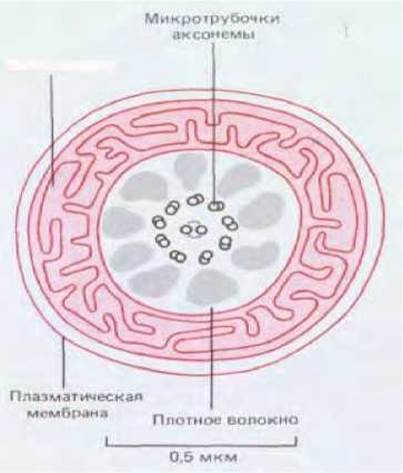
Рис. 15-35. Средняя часть сперматозоида млекопитающего в поперечном разрезе (по данным электронной микроскопии). Жгутик состоит из аксонемы и девяти окружающих ее плотных волокон. Аксонема построена из двух одиночных микротрубочек, окруженных девятью двойными микротрубочками. Расположение митохондрии (изображена в цвете) отлично обеспечивает доставку АТР, необходимого для движения жгутика; ее необычное строение является результатом слияния отдельных митохондрий во время дифференцировки сперматиды
15.3.9. У многих млекопитающих спермии образуются постоянно [16]
Между процессами образования яйцеклеток (оогенез) и сперматозоидов (сперматогенез) у млекопитающих существует ряд важных различий. Например, мы уже говорили о том, что у женщин на ранней стадии эмбриогенеза из размножающихся оогоний образуется ограниченное число ооцитов, которые через определенные промежутки времени по очереди претерпевают овуляцию. С другой стороны, сперматогенез у мужчин начинается только после полового созревания и затем непрерывно продолжается в эпителиальной выстилке очень длинных, сильно извитых трубочек, называемых семенными канальцами, которые находятся в семенниках. Незрелые половые клетки, называемые сперматогониями, располагаются на самой периферии канальца, у базальной мембраны, где они все время делятся путем митоза. Некоторые из дочерних клеток перестают делиться и дифференцируются в сперматоциты первого порядка. Эти клетки вступают в I профазу мейоза, в которой происходит кроссинговер между их спаренными гомологичными хромосомами, а затем заканчивают первое деление, образуя по два сперматоцита второго порядка; у человека каждый из них содержит 22 дуплицированные аутосомы и одну дуплицированную хромосому X или Y. Каждая хромосома по-прежнему состоит из двух сестринских хроматид, и когда оба сперматоцита второго порядка претерпевают второе деление мейоза, образуются четыре сперматиды с гаплоидным числом одиночных хромосом. Затем такие гаплоидные сперматиды в результате морфологической дифференцировки превращаются в зрелые спермии, которые выходят в просвет семенного канальца (рис. 15-37 и 15-38), а позднее - в придаток семенника, представляющий собой извитую трубочку, охватывающую семенник; здесь спермии накапливаются и здесь же продолжается их созревание.
Подобно оогенезу, сперматогенез находится под влиянием гормональной регуляции. После наступления половой зрелости гипофиз мужчины начинает выделять гормон, о котором уже говорилось при обсуждении оогенеза у млекопитающих; это лютеинизирующий гормон (ЛГ). Под воздействием ЛГ клетки Лейдига, расположенные между семенными канальцами в семенниках, выделяют большие количества мужского полового гормона тестостерона. Последний в свою очередь инициирует сперматогенез; возможно, это происходит благодаря действию тестостерона на клетки Сертоли, которые сплошным слоем покрывают развивающиеся спермии, защищая их и обеспечивая их питание (рис. 1538).
Одна из загадочных и уникальных особенностей спермиев состоит в том, что в процессе их развития митотические и мейотические деления не сопровождаются полным, доведенным до конца делением цитоплазмы (цитокинезом); поэтому все дифференцирующиеся дочерние клетки, происходящие из одного сперматогония, соединены цитоплазматическими мостиками (рис. 15-39). Такие мостики остаются до самого конца дифференцировки спермиев, т. е. до того момента, когда отдельные сперматозоиды переходят в просвет канальцев. Группа клеток, связанных подобным образом, называется синцитием. Наличие этой структуры объясняет синхронность появления зрелых спермиев в любом данном участке семенного канальца.
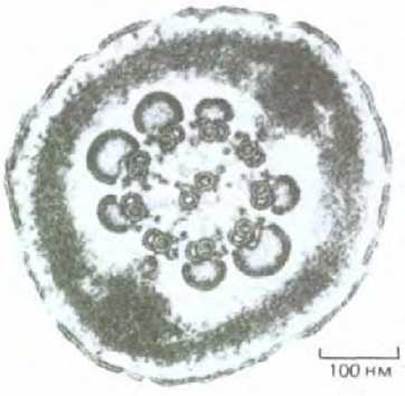
Рис. 15-36. Поперечный срез жгутика сперматозоида морской свинки (электронная микрофотография). Здесь отсутствуют митохондрии, поскольку сечение произведено в области хвоста сперматозоида, позади его средней части. В представленном участке жгутика два из девяти плотных волокон оканчиваются и сливаются с внешней волокнистой оболочкой. (С любезного разрешения Daniel S. Friend.)

Рис. 15-37. Различные стадии сперматогенеза. Сперматогонии развиваются из первичных половых клеток, мигрирующих в семенники на ранней стадии эмбрионального развития. Когда животное достигает половой зрелости, сперматогонии начинают быстро размножаться, причем часть из их потомков сохраняет способность к непрерывным неограниченным делениям (сперматогонии типа стволовых клеток), а другая часть (созревающие сперматогонии) после ограниченного числа последовательных митозов приступает к мейозу, превращаясь в сперматоциты первого порядка. После завершения второго деления мейоза сперматоциты первого порядка превращаются в гаплоидные сперматиды, дифференцирующиеся в зрелые спермии. Сперматогенез отличается от оогенеза (рис. 15-25) в нескольких отношениях: 1) после полового созревания в мейоз непрерывно вступают новые клетки; 2) из каждой приступившей к мейозу клетки образуется не одна, а четыре зрелые гаметы и 3) зрелые спермии формируются после завершения мейоза в ходе сложного процесса клеточной дифференцировки.

Рис. 15-38. Крайне упрощенная схема поперечного разреза семенного канальца из семенника млекопитающего. А. На всех представленных здесь стадиях сперматогенеза развивающиеся гаметы тесно связаны с клетками Сертоли; последние представляют собой крупные клетки, занимающие пространство от базальной мембраны до просвета семенного канальца. На ход сперматогенеза оказывает влияние тестостерон, выделяемый клетками Лейдига, которые располагаются в промежутках между семенными канальцами. Б. Делящиеся сперматогонии находятся вблизи базальной мембраны. Некоторые из этих клеток перестают делиться путем митоза и приступают к мейозу, превращаясь в сперматоциты первого порядка. В конце концов зрелые спермии выходят в просвет канальца. Завершение сперматоцитом мейоза и превращение его в сперматиду занимает у человека около 24 дней, и еще 5 недель требуется на превращение сперматиды в спермий. Спермии претерпевают дальнейшее созревание, обретают подвижность в придатке яичка и лишь тогда становятся совершенно зрелыми.
15.3.10. Ядра спермиев гаплоидны, однако, процессом дифференцировки этих клеток управляет диплоидный геном [17]
В отличие от яйцеклеток у сперматозоидов дифференцировка в основном осуществляется после того, как они завершают мейоз и становятся гаплоидными. Благодаря цитоплазматическим мостикам, каждый развивающийся гаплоидный спермий может получать весь набор продуктов полного диплоидного генома. То, что дифференцировкой спермиев, так же как и дифференцировкой яйцеклеток, управляет диплоидный геном, важно по двум причинам. Во-первых, в исходном диплоидном геноме, как правило, содержится некоторое число дефектных аллелей - рецессивных летальных мутаций (разд. 15.1.4); гаплоидная клетка, получившая один из этих дефектных аллелей, весьма вероятно, погибнет, если она не будет обеспечена продуктами нормального аллеля, закодированными в других ядрах, которые его содержат. Во-вторых, у некоторых организмов (например, у человека) одни спермии получают при мейозе Х-хромосому, а другие - Y-хромосому. Поскольку Х-хромосома содержит много весьма важных генов, отсутствующих в Y-хромосоме, можно думать, что если бы не цитоплазматические мостики между развивающимися спермиями, то те из них, которые получили Y-хромосому, не выжили бы, и в результате в следующем поколении не было бы ни одного мужчины.
И в самом деле, имеются прямые экспериментальные данные о том, что дифференцировкой спермиев управляют продукты диплоидного генома. Часть таких данных была получена при исследовании дефектных в отношении расхождения хромосом мутантов дрозофилы, у которых в процессе мейоза хромосомы неравномерно распределяются между дочерними клетками; в результате одни сперматозоиды содержат слишком мало хромосом, другие - слишком много, а у некоторых их вообще нет. Поразительно то, что дифференцировка всех этих клеток, даже тех, в которых вовсе нет хромосом, протекает нормально. Этот факт можно объяснить на основе упомянутого выше предположения: продукты недостающих хромосом могли бы доставляться путем диффузии по цитоплазматическим мостикам, связывающим соседние клетки. Не исключено и иное объяснение: в диплоидных сперматогониях или сперматоцитах первого порядка могут заранее, еще до мейоза, создаваться «инструкции» для дифференцировки спермия (предположительно в виде долгоживущих мРНК), так что нет необходимости в функционировании гаплоидного генома в период самой дифференцировки. Независимо от того, какое из этих объяснений верно, очевидно, что при дифференцировке спермии используются продукты обоих хромосомных наборов, хотя собственное ядро клетки при этом гаплоидно.
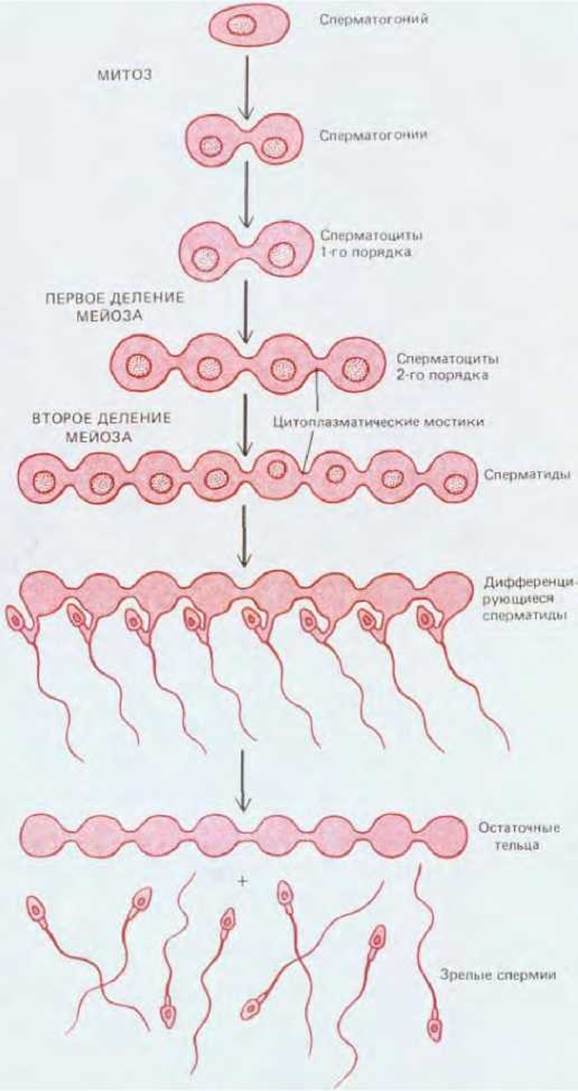
Рис. 15-39. Эта схема показывает, каким образом потомки одного созревающего сперматогония на протяжении всего периода дифференцировки их в зрелые спермии остаются связанными друг с другом цитоплазматическими мостиками. Для простоты показано, что в митоз вступают только два созревающих соединенных сперматогония, из которых в конечном счете образуются восемь связанных между собой гаплоидных сперматид. На самом деле число связанных клеток, проходящих два деления мейоза и совместно дифференцирующихся, значительно больше, чем здесь показано.
Заключение
Яйцеклетки развиваются из первичных половых клеток, которые на ранней стадии развития организма мигрируют в яичник и превращаются там в оогонии. После периода митотического размножения оогонии становятся ооцитами первого порядка, которые, вступив в первое деление мейоза, задерживаются в профазе I на время, измеряемое сутками или годами в зависимости от вида организма. В период этой задержки ооцит растет и накапливает рибосомы, мРНК и белки, зачастую используя при этом другие клетки, включая окружающие вспомогательные клетки. Дальнейшее развитие (созревание яйцеклетки) зависит от полипептидных гормонов (гонадотропинов), которые, воздействуя на окружающие каждый ооцит вспомогательные клетки, побуждают их индуцировать созревание небольшой части ооцитов. Эти ооциты завершают первое деление мейоза, образуя маленькое полярное тельце и крупный ооцит второго порядка, который позже переходит в метафазу второго деления мейоза; у многих видов ооцит задерживается на этой стадии до тех пор, пока оплодотворение не инициирует завершение мейоза и начало развития эмбриона.
Спермий обычно представляет собой маленькую и компактную клетку, которая в высокой степени специализирована для функции внесения своей ДНК в яйцеклетку. В то время как у многих организмов весь пул ооцитов образуется еще на ранней стадии развития самки, у самцов после наступления половой зрелости в мейоз непрерывно вступают все новые и новые половые клетки, причем каждый сперматоцит первого порядка дает начало четырем зрелым спермиям. Дифференцировка спермиев осуществляется после мейоза, когда ядра гаплоидны. Однако, поскольку при митотическом делении зрелых сперматогониев и сперматоцитов цитокинез не доводится до конца, потомки одного сперматогония развиваются в виде синцития. В связи с этим дифференцировка спермия может контролироваться продуктами хромосом от обоих родителей.