Молекулярная биология клетки - Том 3 - Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. 1994
От клеток к многоклеточным организмам
Клеточные механизмы развития
Морфогенетические движения и формирование общей пространственной организации тела
В этом и следующих разделах речь пойдет о том, как возникает пространственная организация раннего эмбриона и какие физические силы участвуют в его формировании. В качестве примера мы в основном будем обращаться к шпорцевой лягушке Xenopus laevis (рис. 16-1), раннее развитие которой было изучено особенно тщательно. Подобно зародышам других земноводных, зародыши Xenopus относительно устойчивы к внешним воздействиям и поэтому служат удобным объектом для экспериментов.
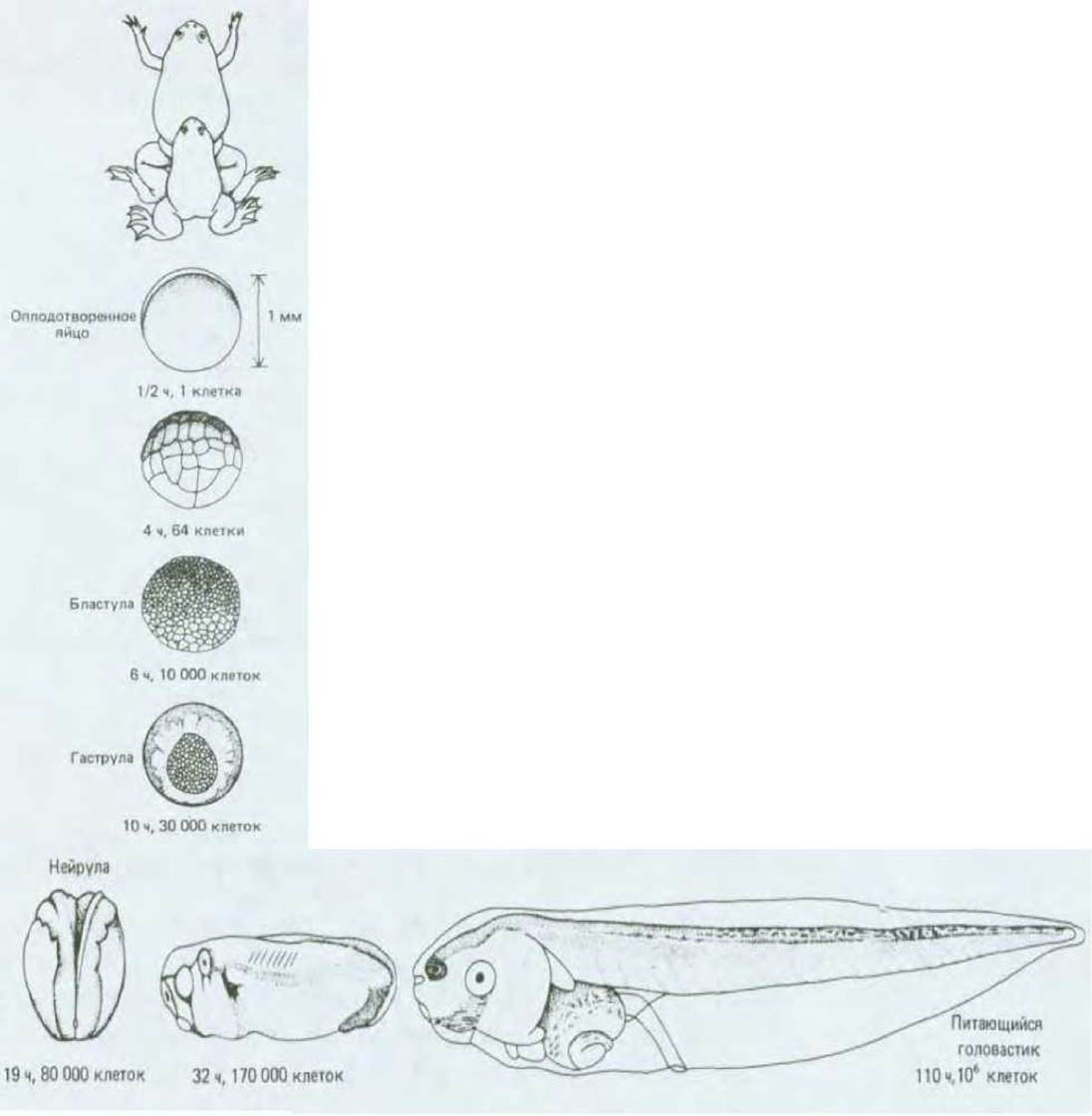
Рис. 16-1. Схема развития Xenopus laevis от оплодотворенного яйца до самостоятельно питающегося головастика. Вверху - спаривание взрослых самца и самки. Ниже представлены последовательные стадии развития (ранние стадии - вид сбоку; 10-часовой эмбрион-вид снизу; 19- часовой-вид сверху). Все рисунки (кроме верхнего) выполненны в одном масштабе. [P. D. Nieuwkoop, J. Faber, Normal Table of Xenopus laevis (Daudin), Amsterdam: North-Holland, 1956.]
Для того чтобы облегчить восприятие излагаемого материала, процесс эмбриогенеза позвоночных (и многих других групп животных) подразделен на три периода. Первый - это дробление оплодотворенного яйца на множество более мелких клеток, формирующих слой наподобие эпителия, из которого в результате процессов гаструляции и нейруляции образуются полость первичной кишки и нервная трубка. Затем следует период органогенеза, т. е. возникают различные органы и части тела (конечности, глаза, сердце и т. д.). Третий период развития характеризуется тем, что органы, сформировавшиеся в виде небольших структур, растут, пока не достигнут размеров, свойственных взрослому животному. Эти периоды не имеют четких границ и могут в значительной степени перекрываться. Развитие от стадии оплодотворенного яйца до начала органогенеза мы проследим на примере эмбриогенеза у Хеnорus.
16.1.1. Полярность эмбриона земноводных определяется полярностью яйца [2]
Яйцо земноводных - это сравнительно крупная клетка (около 1 мм в диаметре), покрытая прозрачной неклеточной капсулой - студенистой оболочкой. Большая часть клетки заполнена желточными пластинками, состоящими в основном из белка и липидов. Желток сконцентрирован в нижней половине яйца, называемой вегетативным полушарием; верхнюю половину яйца называют анимальным полушарием. Оплодотворение инициирует превращение этой единичной клетки в многоклеточную личинку (головастика), у которой должны определяться передний и задний концы тела, брюшная и спинная стороны, а также срединная плоскость симметрии, разделяющая тело на правую и левую половины. Развитие зародыша удобно описывать, пользуясь системой трех осей: переднезадней (от головы к хвосту), дорсовентральной (от спины к животу) и медиолатеральной (от срединной плоскости влево и вправо). Такая полярность зародыша закладывается у земноводных на очень раннем этапе развития. Еще до оплодотворения анимальный и вегетативный полюсы яйца содержат различные наборы мРНК, равно как и разное количество белка и других клеточных компонентов, но при этом яйцо симметрично относительно анимально- вегетативной оси. Эта исходная анимально-вегетативная симметрия достаточна для создания цилиндрического эмбриона, обладающего переднезадней осью.
Оплодотворение обеспечивает углубление различий в содержимом яйца земноводных, реализуемое в появлении дорсовентральной полярности. Наружный обогащенный актином слой цитоплазмы, называемый кортикальным слоем, или кортексом, внезапно поворачивается по отношению к внутреннему содержимому как единый сегмент, так что анимальный полюс кортекса смещается по отношению к анимальному полюсу внутреннего содержимого в направлении будущей вентральной стороны (рис. 16-2). Направление вращения определяется местом проникновения спермия; объяснить это, по-видимому, можно влиянием центросомы, привносимой спермием в яйцо. Поскольку такое вращение вызывает в яйце смещение пигментных гранул, у некоторых земноводных появляется слабо пигментированная полоска, называемая серым серпом. Она образуется на стороне яйца, противоположной месту проникновения спермия. Место проникновения спермия соответствует, грубо говоря, брюшку; на противоположной стороне формируются дорсальные структуры, например спинной мозг (рис. 16-3). В результате воздействий, блокирующих вращение кортекса, развиваются так называемые радиализованные животные с кишечником, расположенным в центре тела, у которых отсутствует дорсовентральная асимметрия.
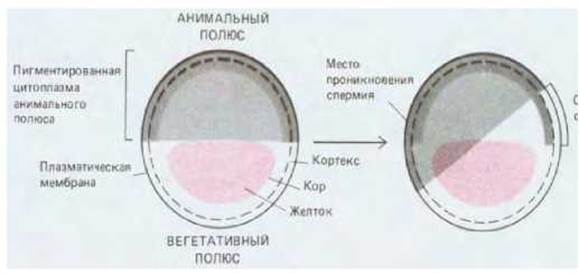
Рис. 16-2. Первое морфогенетическое движение после оплодотворения яйца лягушки. Кортекс яйца слой, толщиной примерно в несколько микрометров - поворачивается почти на 30° относительно внутренних слоев яйца в направлении, которое определяется местом проникновения спермия. У видов с окрашенной цитоплазмой анимального полюса такое вращение приводит к появлению четко наблюдаемого серого серпа, который расположен напротив места проникновения спермия.
16.1.2. В результате дробления из одной клетки образуется множество клеток [3]
Поворот кортикального слоя завершается примерно через час после оплодотворения и создает условия для осуществления дробления, в ходе которого из одной крупной яйцеклетки за счет повторяющихся митозов образуется множество более мелких клеток -бластомеров; общая масса эмбриона при этом не меняется. Чтобы выжить, эмбрион должен быстро достичь стадии, когда он сможет самостоятельно питаться, плавать и спасаться от хищников, и поэтому первые деления дробления очень быстро следуют друг за другом; в этот период длительность клеточного цикла составляет около 30 мин (см. разд. 13.1.9). Высокая скорость репликации ДНК и чередования митозов не позволяет осуществлять транскрипцию генов и дробящийся эмбрион полностью зависит от запасов РНК, белка, мембран и других материалов, которые были накоплены в яйце в процессе его созревания в материнском организме. Только биосинтез ДНК имеет теперь жизненно важное значение, и ее необычно быстрая репликация становится возможной благодаря огромному числу мест, в которых начинается этот процесс (см. разд. 9.3.9). Борозда первого деления дробления делит яйцо по вертикали, т. е. в плоскости, проходящей через анимально-вегетативную ось; в результате образуются две симметричные половины яйца (рис. 16-4). Следующее деление вновь происходит вертикально, но под прямым углом к плоскости первого деления и приводит к образованию четырех клеток одинаковой величины. Борозда третьего деления дробления располагается горизонтально несколько выше срединной плоскости, поэтому образующиеся четыре верхние клетки меньше по размерам, чем нижние; к тому же нижние клетки содержат больше желтка. Примерно после 12 клеточных циклов скорость деления клеток резко снижается, нарушается также синхронность делений и начинается транскрипция генома зародыша. Это изменение, известное как переход к средней бластуле (ПСБ), по всей вероятности, отражает истощение определенного клеточного компонента материнского происхождения, который используется для связывания с вновь синтезируемой ДНК.
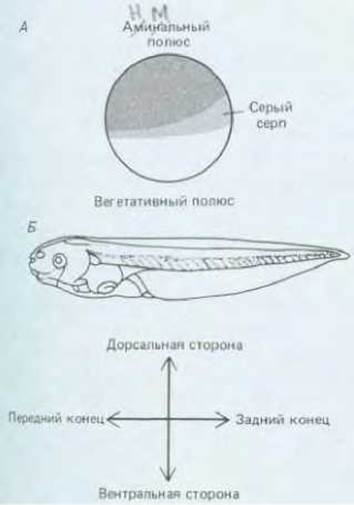
Рис. 16-3. Асимметрия яйца Хепорus показана на рис. А. Она определяет расположение осей тела будущего головастика, изображенное на рис. Б. Точная привязка осей яйца к телу головастика сложна, поскольку гаструляционные движения значительно меняют топологию зародыша. В первом приближении анимальный полюс соответствует переднему концу тела (голове), а серый серп соответствует дорсальной стороне тела.
У других животных взаимная ориентация плоскостей последовательных делений дробления может быть иной. В яйцах очень богатых желтком как, например, у птиц, борозды делений дробления не в состоянии разбить желток на части, и поэтому все ядра сосредоточены на анимальном полюсе; в результате зародыш развивается только из «шапочки» клеток, лежащих поверх желтка.
16.1.3. Бластула представляет собой полый шар, стенка которого образована одним слоем клеток [4]
Уже в самом начале развития эмбриона его клетки связаны между собой не только механически, но и с помощью щелевых контактов, через которые способны проходить ионы и другие низкомолекулярные вещества, передавая «послания», которые могут играть важную роль на более поздних стадиях развития (вместе с другими типами сигналов) для координации поведения клеток (см. разд. 14.1.7). На периферии зародыша бластомеры связаны друг с другом плотными контактами (см. разд. 14.1.1); это позволяет изолировать внутреннюю часть зародыша от окружающей среды. Примерно на стадии 16 бластомеров промежутки между клетками в центральной части зародыша расширяются и образуют единую полость- бластоцель; это результат переноса ионов натрия во внутренние межклеточные пространства через мембраны клеток; осмотическое давление внутри зародыша повышается и сюда начинает поступать вода. Клетки, окружающие бластоцель, образуют эпителиоподобный слой; теперь эту стадию развития называют бластулой (рис. 16-5). То, что клетки бластулы организованы как эпителиальный слой, жизненно важно для координации их дальнейшего поведения.
16.1.4. После гаструляции полый клеточный шар превращается в трехслойную структуру с первичной кишкой [5, 6]
После того как клетки бластулы сформировали эпителиальный слой1, наступает время для координированных движений, приводящих к образованию гаструлы. Процесс образования гаструлы называют гаструляцией. Эта радикальная перестройка ведет к превращению полого клеточного шара в многослойную двустороннесимметричную структуру с кишечной трубкой, располагающейся в центре; в результате сложного процесса инвагинации (впячивания) значительный участок эпителия перемещается с наружной поверхности внутрь зародыша. Последующее развитие теперь уже определяется взаимодействиями внутреннего, наружного и среднего слоев, возникающих в результате гаструляции. Гаструляция в той или иной форме происходит у всех многоклеточных животных. Так как с геометрической точки зрения гаструляция у земноводных выглядит «искаженной», мы сначала рассмотрим этот процесс у морского ежа - близкого родственника позвоночных (см. рис. 15-40, разд. 15.4.1).
1 Автор в данном случае употребляет не совсем точный термин, называя стенку бластулы эпителием или эпителиальным слоем. Дело в том, что эпителий - это дифференцированная ткань, образующаяся на более поздних стадиях развития из эктодермы (наружного зародышевого листка). Стенку зародыша на стадии бластулы называют бластодермой, и она лишь чисто внешне напоминает эпителиальную ткань. Из чисто технических соображений мы не во всех случаях исправляли эту неточность, и читателю следует иметь это в виду. - Прим. ред.

Рис. 16-4. Стадии дробления у Xenopus (вид сбоку).

Рис. 16-5. Бластула. На этой стадии развития клетки образуют эпителиоподобный слой, окружающий полость, заполненную жидкостью,-бластоцель. Щелевые контакты обеспечивают электрическое сопряжение клеток, а плотные контакты наружной поверхности изолируют внутреннее пространство эмбриона от окружающей среды. Обратите внимание, что у Xenopus стенка бластулы образована несколькими слоями клеток, и только наружные клетки плотно прилегают друг к другу, как это имеет место в любом типе эпителия.
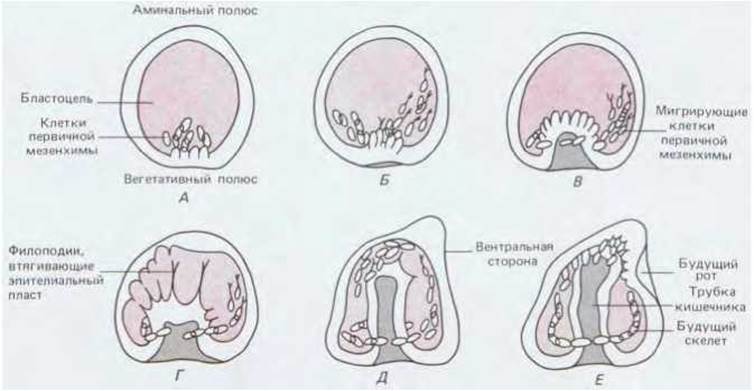
Рис. 16-6. Гаструляция у морского ежа. На вегетативном полюсе бластулы от ее стенки отделяются клетки первичной мезенхимы (А). Эти клетки «ползут» вверх по внутренней поверхности стенки бластулы (Б). В это время стенка бластулы в области вегетативного полюса (вегетативная пластинка) начинает впячиваться внутрь (В). Клетки на верхушке инвагинирующей вегетативной пластинки образуют филоподии, прикрепляющиеся к поверхности внутренней стенки бластоцеля и подтягивают ее дальше в глубь бластоцеля, образуя полость первичной кишки (Г, Д). Конец кишечной трубки вступает в контакт со стенкой бластулы; в этом месте впоследствии возникает ротовое отверстие (Е). (L. Wolpert, T. Gustafson, Endeavour, 26, 85-90, 1967.)
Эмбрион морского ежа прозрачен, поэтому развитие его, внешнее и внутреннее, можно исследовать прижизненно; в то же время на этом объекте удобно изучать активность отдельных клеток. Исходным материалом для гаструляции является бластула, устроенная довольно просто: это - полый шар, стенку которого образуют около 1000 клеток, расположенных в один слой; полость бластулы называют бластоцелем. Всю эту структуру покрывает тонкий слой внеклеточного матрикса; у бластулы можно также различать вегетативный и анимальный полюсы. Гаструляция начинается с того, что на вегетативном полюсе от эпителия отделяются несколько десятков так называемых клеток первичной мезенхимы (рис. 166, А). По всей вероятности, эти клетки теряют способность связываться с другими клетками и внеклеточным матриксом наружной поверхности эмбриона и приобретают сродство к обогащенному фибронектином матриксу (см. разд. 14.2.13), который выстилает бластоцель. Эти клетки выходят в полость бластулы и движутся вдоль ее стенки, подтягиваясь на выпускаемых ими длинных тонких отростках (филоподиях) с «липкими» концами (рис. 16-7). Когда кончик филоподии вступает в контакт с поверхностью, к которой он может плотно прикрепиться, филоподия сокращается и тянет за собой клетку. Образовавшиеся филоподии, по-видимому, втягиваются обратно, а вместо них в других местах возникают новые, так что клетка может перемещаться то в одном, то в другом направлении. В конце концов, однако, клетки занимают четко определенное положение, что, по всей вероятности, обусловлено их специфическим сродством к каким-то участкам поверхности бластоцеля. Это было показано в опытах с моноклональными антителами, которые продемонстрировали высокоспециализированные различия между клетками в разных участках эмбриона в отношении химизма их поверхности (к этой теме мы вернемся позже - см. разд. 16.6.1). Коль скоро клетки первичной мезенхимы заняли свое место, они начинают формировать скелет (16-6, Е).
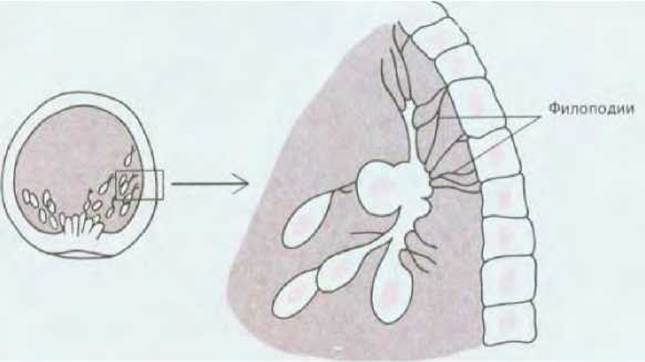
Рис. 16-7. Клетки первичной мезенхимы ползут по внутренней поверхности стенки бластулы, выпуская сократимые филоподии с «липкими» концами. (L. Wolpert, Т. Gustafson, Endeavour, 26, 85-90, 1967.)
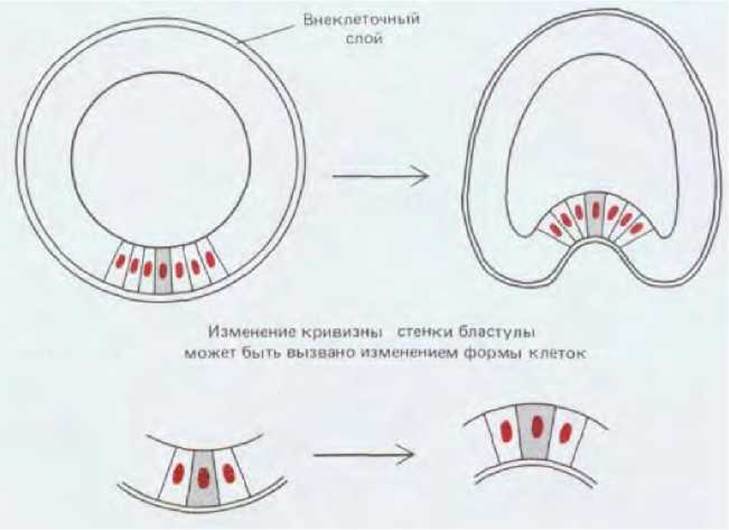
Рис. 16-8. Эта схема иллюстрирует возможный механизм начальных этапов впячивания клеток на вегетативном полюсе. Детали этого процесса на молекулярном уровне рассмотрены в разд. 16.1.6 и 11.6.9.
С началом миграции клеток первичной мезенхимы начинает инвагинировать (впячиваться) эпителий в области вегетативного полюса, образуя, таким образом, первичную кишку (рис. 16-6, В). При этом сначала изменяется форма эпителиальных клеток: внутренний конец клетки, обращенный к бластоцелю, становится шире чем наружный, и поэтому клеточный слой прогибается внутрь бластоцеля (рис. 16-8). Следующий этап инвагинации происходит за счет иного процесса-перераспределения клеток. Инвагинирующие клетки активно перестраиваются, но их форма при этом не меняется. В результате поначалу довольно широкая полость гаструлы превращается в длинную узкую кишечную трубку. Одновременно определенные клетки на верхушке этой кишечной трубки выпускают в бластоцель длинные филоподии, которые вступают в контакт со стенками полости, прилипают к ним и сокращаются, как бы помогая направлять процесс инвагинации (рис. 16-6, Г, Д). Движение прекращается после соприкосновения слепого конца кишечной трубки с наружной стенкой зародыша на его противоположном конце (рис. 16-6, Е). Позднее в месте контакта двух соприкасающихся слоев стенка зародыша прорывается, и на месте прорыва образуется вторичный рот. Поскольку клетки, которые своими филоподиями направляли инвагинацию, выполнили свою задачу, они отделяются от эпителия, перемещаются в пространство между кишечной трубкой и стенкой тела и превращаются в так называемую вторичную мезенхиму, которая со временем даст начало стенке целома и мускулатуре.
В результате гаструляции полая сферическая бластула превращается в трехслойную структуру: внутренний слой, т.е. стенку первичной кишки, называют энтодермой; наружный слой, который так и остался снаружи, - эктодермой, а промежуточный рыхлый слой ткани, состоящий из первичной и вторичной мезенхимы, - мезодермой. Это - три первичных зародышевых листка, характерные для всех высших животных. Организация трехслойного эмбриона в общих чертах соответствует организации взрослого животного с пищеварительной трубкой внутри, эпидермисом снаружи и органами соединительного тканного происхождения между ними. В первом приближении можно сказать, что эти три типа тканей взрослого организма происходят соответственно из энтодермы, эктодермы и мезодермы, хотя встречаются и исключения (см. разд. 16.1.7, 16.1.8 и 16.1.9).
16.1.5. Гаструляционные движения основаны на четко скоординированных простых движениях клеток [1, 6, 7]
Гаструляционные движения основаны на относительно простом наборе основных движений клеток. Клетки могут менять свою форму в результате вытягивания или сокращения; они приклеиваются или отделяются от других клеток или внеклеточного матрикса; они могут секретировать вещества внеклеточного матрикса, которые сдерживают или направляют их движения. Такие виды клеточной активности, наряду с ростом клеток и их делением, лежат в основе всех морфогенетических движений как отдельных клеток, так и их групп, включая, в частности, такие процессы, как перераспределение клеток, обеспечивающее инвагинацию и образование первичной кишки. Гаструляция, будучи исключительно важным событием per se, кроме того, демонстрирует различные формы клеточного поведения.
В последующих главах будут обсуждаться механизмы, используемые клетками для изменения своей формы или адгезивных свойств. Индивидуальное развитие ставит перед нами особую проблему: необходимо понять, каким образом элементарные виды клеточной активности координированы во времени и пространстве; именно эта координация определяет поведение каждой из частей эмбриона. В настоящее время нам еще не вполне ясны прекрасно срежиссированные гаструляционные движения. Тем не менее эксперименты на земноводных позволяют ответить на вопрос, какие из частей гаструлирующего эмбриона движутся за счет своих собственных возможностей и какие части эмбриона находятся под воздействием внешних сил.
У зародышей земноводных присутствие клеток со значительным избытком желтка существенно замедляет инвагинацию, вследствие чего геометрия процесса становится более сложной, чем у морского ежа. Инвагинация начинается не на вегетативном полюсе, а несколько сбоку, где сначала образуется небольшое углубление, именуемое бластопором. Через него происходит вворачивание клеток будущей энтодермы (рис. 16-9). По мере углубления процесса инвагинации бластопор приобретает форму кольца, окружающего часть видимой снаружи энтодермы. Эта часть энтодермы называется желточной пробкой; здесь расположены клетки, обогащенные желтком (позже они попадут в полость кишки и будут переварены). В это же время бластодерма на анимальном полюсе активно расширяется и занимает место клеток, устремляющихся внутрь. В конце концов бластодерма анимального полушария покрывает всю наружную поверхность эмбриона, а размеры бластопора сильно уменьшаются.
В основе процесса инвагинации у земноводных, по-видимому, лежат те же механизмы, что и у морского ежа. Сначала происходит изменение формы клеток в области бластопора. У земноводных это так называемые колбовидные клетки; они характеризуются широким основанием и длинным узким концом, прикрепляющим клетки к внутренней поверхности стенки бластулы (рис. 16-10). Эти клетки тянут эпителий внутрь, вследствие чего на поверхности бластулы возникает продольная борозда. После формирования этой борозды клетки продолжают свое движение внутрь в виде пласта, принимающего участие в формировании кишки. Как и у морского ежа, это движение осуществляется за счет активной переупаковки клеток эпителия, особенно в краевой зоне, прилежащей к губе бластопора (рис. 16-10). Небольшие квадратные фрагменты изолированной ткани краевой зоны в условиях культуры могут спонтанно удлиняться и расширяться вследствие переупаковки клеток (рис. 16-10, Б), как если бы они находились в зародыше и устремлялись внутрь последнего через бластопор. Механизм перераспределения клеток, лежащий в основе этого конвергентного расширения остается загадкой. Вторичная направляющая, или движущая, сила обеспечивается будущими клетками мезодермы, которые мигрируют по внутренней поверхности крыши бластоцеля и тянут энтодерму за собой, подобно тому как это происходит у зародыша морского ежа, только у морского ежа в качестве движущей силы выступают будущие клетки вторичной мезенхимы. Крыша бластоцеля выстлана матриксом, богатым фибронектином, и при инъекции в бластоцель антител или пептидов, блокирующих взаимодействие фибронектина с рецепторами клеточной поверхности (см. разд. 14.2.13), нормальная гаструляция тормозится (в частности, у тритонов и саламандр) (рис. 16-11).
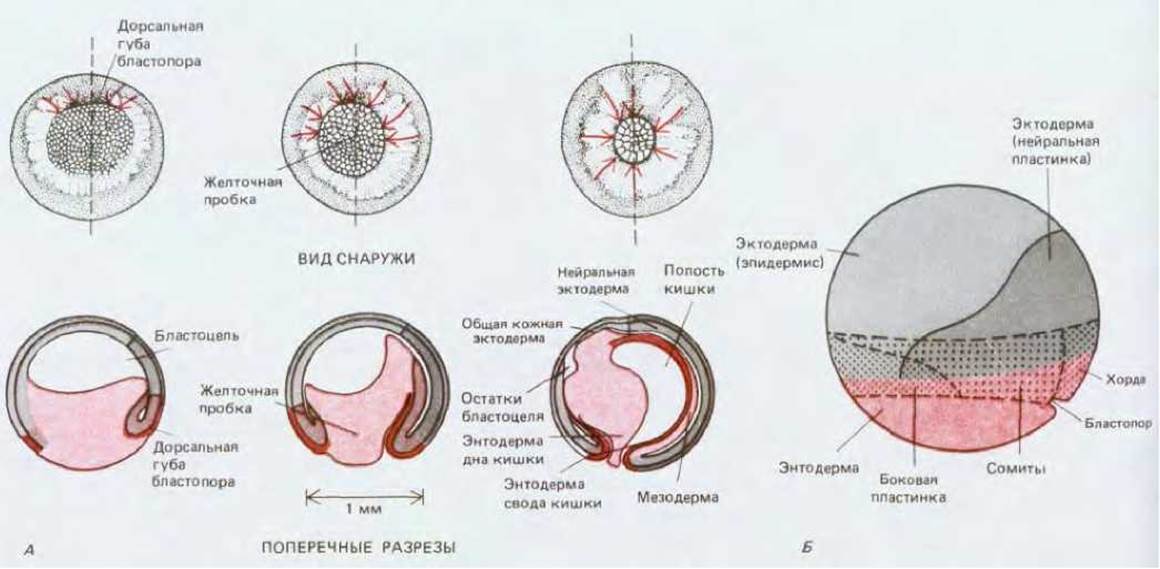
Рис. 16-9. А. Гаструляция у Xenopus. Внешний вид эмбриона (вверху) представлен со стороны вегетативного полюса; поперечные разрезы (внизу) проходят в плоскости, показанной штриховой линией. Направление движения клеток указано стрелками. Б. Карта презумптивных зачатков раннего эмбриона Xenopus (вид сбоку) с начала гаструляции указывает на происхождение клеток, которые вследствие гаструляционных движений примут участие в формировании трех зародышевых листков. Различные части мезодермы (боковая пластинка, сомиты, хорда) происходят из более глубоко расположенных клеток области, обозначенной точками; другие клетки этой области, в том числе наиболее поверхностные, дадут начало эктодерме (серый цвет, вверху) или энтодерме (различные оттенки красного цвета, книзу). (R. Е. Keller, J. Exp. Zoob, 216, 81-101, 1981.)
У земноводных, как и у морских ежей, гаструляция приводит к образованию трехслойного зародыша с наружным эктодермальным слоем, внутренней трубкой энтодермы, представляющей собой зачаток кишки, и мезодермой между ними. Ротовое отверстие образуется на переднем конце зародыша, где мезодерма отсутствует, и поэтому эктодерма может вступать в прямой контакт с энтодермой.

Рис. 16-10. А. На срезе через эмбрион Xenopus, сделанном во время гаструляции и проходящем в той же плоскости, что и на рис. 16-9, показаны четыре основных типа движений, лежащих в основе этого процесса. Б. Модель перераспределения клеток, приводящего к конвергентному вытягиванию, которое, возможно, является главной движущей силой процессов гаструляции у Xenopus. (А - из R.E. teller, J. Exp. Zool., 216, 81-101, 1981; Б-т J. Gerhart, R. Keller, Annu. Rev. Cell Biol., 2, 201 229, 1986.)
16.1.6. Гаструляционные движения организованы вокруг бластопора [2, 7, 8]
Движения клеток в процессе гаструляции сложны, но упорядочены, что позволяет еще до гаструляции построить на поверхности эмбриона карту презумптивных зачатков (рис. 16-9, Б), которая показывает, из каких клеток предстоит формироваться различным частям взрослого организма. А как запускается и организуется весь сложный комплекс гаструляционных движений? У земноводных важным предварительным этапом является перераспределение содержимого яйца сразу же после оплодотворения. Инвагинация всегда начинается в том месте, которое соответствует расположению серого серпа (см. разд. 16.1.1): здесь в области будущей дорсальной губы бластопора вращение кортекса яйца по отношению к внутреннему содержимому, вызванное оплодотворением (см. рис. 16-2), по всей вероятности, создает сочетание клеточных компонентов, обладающее уникальными свойствами. Если в начале гаструляции у зародыша-донора удалить дорсальную губу бластопора и пересадить ее другому зародышу, то гаструляция зародыша - реципиента начнется как в области собственной губы бластопора, так и в области, куда была пересажена губа донора (рис. 16-12). Вследствие этой второй гаструляции происходит образование второго набора структур тела и возникает двойной зародыш (наподобие сиамских близнецов). Если для опытов с пересадкой использовать клетки, которые пигментированы иначе, то ткань реципиента легко отличить от имплантированной ткани. С помощью этого метода удалось показать, что имплантированная губа бластопора вовлекает в контролируемый ею процесс инвагинации эпителий хозяина. Далее мы покажем, что для образования трех зародышевых листков в процессе гаструляции исключительно важны химические и физические взаимодействия между клетками. Но сперва мы вкратце рассмотрим дальнейшее развитие энтодермы, мезодермы и эктодермы, из которых состоит эмбрион позвоночных после окончания гаструляции.

Рис. 16-11. А. Нормальный эмбрион тритона в конце гаструляции; энтодерма и мезодерма инвагинировали внутрь, бластопор приобрел форму кольца, окружающего крупные энтодермальные клетки, еще остающиеся на поверхности. Этот оставшийся участок энтодермы называют желточной пробкой. Б. Такой же эмбрион, у которого гаструляционные движения были блокированы инъекцией в полость бластулы антител против фибронектина. Клетки вегетативного полушария не способны перемещаться внутрь и остаются снаружи (гладкая округлая масса в нижней части рисунка); клетки анимального полушария образуют структуру, напоминающую пустой мешок, и вследствие гаструляционных движений, которые в норме приводят к его распространению по поверхности эмбриона, он стягивается в складки. [J. С. Boucaut et al., J. Embryol. Exp. Morphol., (Suppl.), 211-227, 1985.]
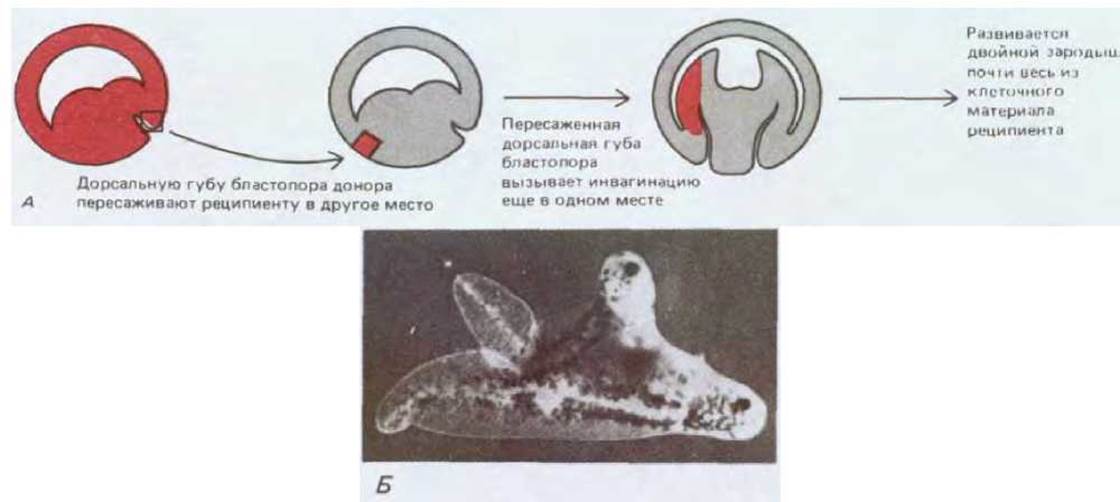
Рис. 16-12. А. Схема эксперимента, показывающего, что дорсальная губа бластопора инициирует и контролирует гаструляционные движения и в случае пересадки организует второй набор структур зародыша. Б. Фотография возникающего в результате такой операции аксолотля, имеющего две головы и два хвоста. Подобные результаты получены и на Xenopus, хотя масштабы дупликации несколько отличаются. (Б-с любезного разрешения Jonathan Slack.)
16.1.7. Из энтодермы образуется кишка и такие ее производные, как легкие и печень [9]
Энтодерма образует трубку-зачаток пищеварительного тракта; она тянется от рта до ануса. Эта трубка дает начало не только глотке, пищеводу, желудку и кишечнику, но также многим железам: слюнным, печени, поджелудочной железе; трахея и легкие также формируются из выростов стенки пищеварительного тракта, который вначале устроен весьма просто. Все эти выросты увеличиваются и превращаются в систему разветвленных трубочек, впадающих в кишку или гортань. Если говорить точнее, энтодерма формирует только внутренние эпителиальные компоненты этих структур - выстилку кишечника и секреторные клетки поджелудочной железы, а поддерживающие их мышечные или соединительнотканные элементы возникают из мезодермы.
16.1.8. Из мезодермы образуются соединительная ткань и мышцы, а также сердечно-сосудистая и мочеполовая системы [9, 10]
После завершения гаструляции образовавшийся средний зародышевый листок, мезодерма, разделяется на две части - для правой и для левой половины тела. Один из участков мезодермы в этот период специализируется, располагаясь вдоль центральной оси тела, и детерминирует деление тела на две половины - правую и левую. Это-хорда, представляющая собой тонкий клеточный тяж диаметром 80 мкм; над ней расположена эктодерма, под ней - энтодерма, а по бокам - мезодерма (см. рис. 16-15). Клетки хорды набухают, в них появляются вакуоли; это приводит к удлинению хорды и как следствие - к выпрямлению зародыша. У самых примитивных хордовых, не имеющих позвоночника, хорда выполняет функции опорной структуры. У позвоночных хорда является той осью, вокруг которой собираются мезодермальные клетки и образуют позвоночник. Таким образом, хорда служит предшественником позвоночного столба не только в процессе эволюции, но и в онтогенезе.
Мезодерма в основном дает начало соединительным тканям - сначала мезенхиме, клетки которой образуют рыхлую сеть, заполняющую промежутки между другими тканями (см. рис. 14-20), а затем кости, хрящу, мышцам и фиброзным тканям, в том числе внутреннему слою кожи (дерме). Мышечные клетки также возникают из мезодермы. Из нее формируется большая часть протоков мочеполовой системы, а также сердечнососудистая система, включая сердце и клетки крови.
16.1.9. Из эктодермы образуется эпидермис и нервная система [9, 11]
Наружная стенка зародыша после гаструляции представлена листком эктодермы, из которой позже образуется наружный слой кожи - эпидермис. Однако этим будущее эктодермы не исчерпывается: из эктодермы формируется также вся нервная система. Процесс образования этой системы называется нейруляцией; он начинается с утолщения широкого дорсального участка эктодермы, который затем сворачивается в трубку и отделяется от остальной части клеточного слоя. Такое преобразование индуцируется хордой и мезодермой, лежащими под этой областью эктодермы (см. разд. 16.6.3). Трубка, образовавшаяся из эктодермы, носит название нервной трубки; в процессе дальнейшего развития из нее возникнет головной и спинной мозг. Вдоль линии, по которой нервная трубка отделяется от будущего эпидермиса, от него обособляется еще некоторое число эктодермальных клеток; позже эти клетки поодиночке мигрируют через мезодерму. Это клетки нервного гребня, из которых образуются практически все компоненты периферической нервной системы (в том числе сенсорные и симпатические ганглии, шванновские клетки, образующие миелиновую оболочку периферических нервов) (см. разд. 19.1.1), а также клетки надпочечников, секретирующие адреналин, и пигментные клетки кожи. В области головы многие клетки нервного гребня дифференцируются в хрящ, кость и иные типы соединительной ткани, которые в других частях тела формируются из мезодермы. Это один из немногих случаев, не укладывающийся в общее представление о соответствии трех зародышевых листков трем «концентрическим» слоям взрослого организма. Органы чувств, передающие нервной системе информацию о зрительных, звуковых, обонятельных и иных стимулах, также развиваются из эктодермальных закладок - одни из нервной трубки, другие - из нервного гребня, третьи - из наружного слоя эктодермы (см. рис. 19-55, разд. 19.7.1). Например, сетчатка образуется как вырост мозга и, следовательно, является производным нервной трубки, тогда как обонятельные клетки дифференцируются прямо из эктодермального эпителия носовой полости.
16.1.10. Нервная трубка образуется в результате координированных изменений формы клеток [6, 12]
Образование нервной трубки - это событие, весьма достойное нашего внимания (рис. 16-13). Сначала поверхность гаструлы представляется довольно однородной, но некоторые изменения здесь уже происходят: эктодерма возле средней линии начинает приподниматься, образуя нервную пластинку. Боковые края нервной пластинки утолщаются; эти утолщения, называемые нервными валиками, постепенно сближаются, а в самой пластинке вдоль средней линии образуется желобок. В конце концов валики сходятся над ним и сливаются, в результате чего возникает полая нервная трубка, покрытая сверху сплошным слоем эктодермы. Как и в случае гаструляции, все эти процессы обусловлены вытягиванием, адгезией и сокращением отдельных клеток эпителиального слоя. Клетки нервной пластинки связаны между собой прочными латеральными соединениями. Вначале клетки удлиняются в направлении, перпендикулярном клеточному слою. Такое удлинение формы клеток связано с удлинением микротрубочек и необходимо для продолжения нейруляции: при воздействии колхицина - вещества, вызывающего разрушение микротрубочек, нервные валики не образуются вовсе или же - коль скоро они возникли - исчезают. Удлинившиеся клетки приобретают затем клиновидную форму с узкими концами, направленными к верхней (апикальной) поверхности клеточного слоя. Так как клетки прочно соединены между собой боковыми поверхностями, а ширина их у основания остается прежней, клеточный пласт в целом прогибается (рис. 16-14). Сужение клеток на верхушке происходит вследствие сокращения пучков актиновых филаментов, которые проходят под апикальной поверхностью клеток, там, где клетки соединяются поясами адгезии (см. разд. 14.1.3).
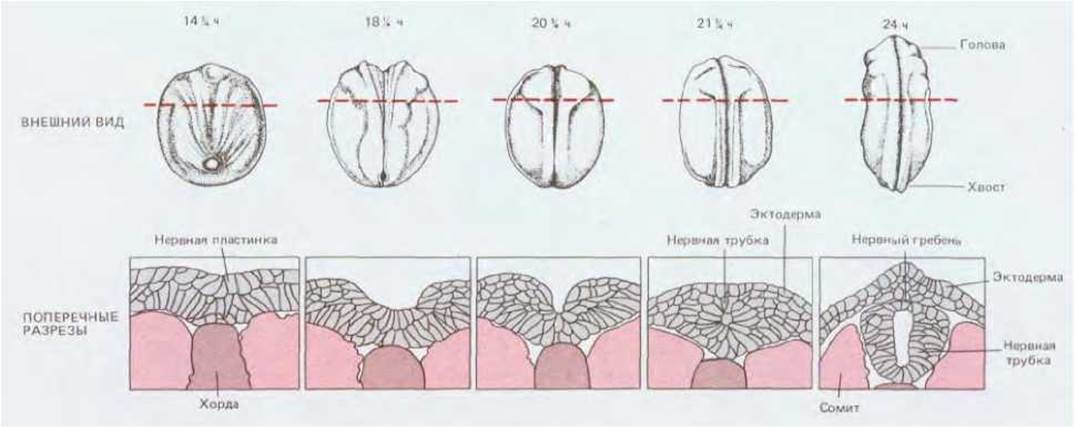
Рис. 16-13. Образование нервной трубки у Xenopus. Вид снаружи с дорсальной стороны. Разрезы проходят в плоскости, указанной прерывистой линией. (Т. Е. Schroeder, J. Embryol, Exp. Morphol., 23, 427-462, 1970.)
16.1.11. Скопление клеток мезодермы делится, образуя сомиты по обе стороны от продольной оси тела [13]
По бокам от вновь образованной нервной трубки лежат обширные участки мезодермы (рис. 16-15). Из утолщенной медиальной области мезодермы образуются позвонки, ребра, скелетные мышцы, а также соединительнотканный слой кожи. Вначале мезодерма на каждой из сторон представлена единой массой ткани, но вскоре она делится на «блоки», называемые сомитами (рис. 16-16). Каждый сомит соответствует одному элементу в возникающем в результате дифференцировки скелете и будет делиться впоследствии на три части. Часть сомита, обращенная к хорде, называется склеротомом и является источником клеток, образующих ребра и позвонки; часть, обращенная наружу и прилежащая к дорсальной поверхности эмбриона, называется дерматомом и является источником клеток, формирующих соединительнотканный слой кожи, или дерму; оставшаяся часть сомита именуется миотомом (между склеротомом и дерматомом) и является источником клеток скелетных мышц.
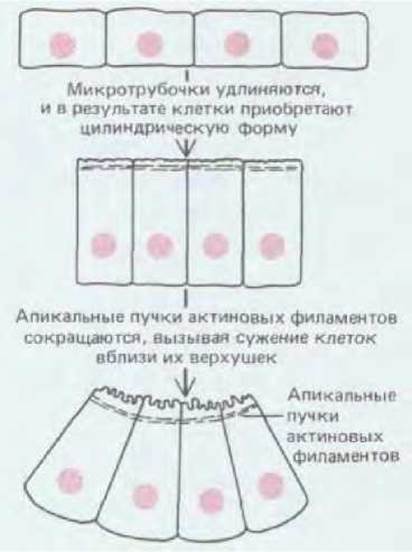
Рис. 16-14. Изгибание клеточного пласта вследствие изменений формы клеток, опосредованных взаимодействием микротрубочек и актиновых филаментов. По мере сужения апикальных концов клеток мембраны их наружных поверхностей становятся вогнутыми. Это может являться симптомом быстрого изменения формы вследствие сокращений, которые имеют место в условиях плотного соприкосновения боковых поверхностей соседних клеток, где тормозится перетекание избытка мембраны вдоль контактирующих комплексов по бокам клеток.

Рис. 16-15. Схематическое изображение поперечного разреза через туловищную область эмбриона лягушки после замыкания нервной трубки. (Т. Mohun, R. Tilly, R. Mohun and J. M. W. Slack, Cell, 22, 9-15, 1980.)
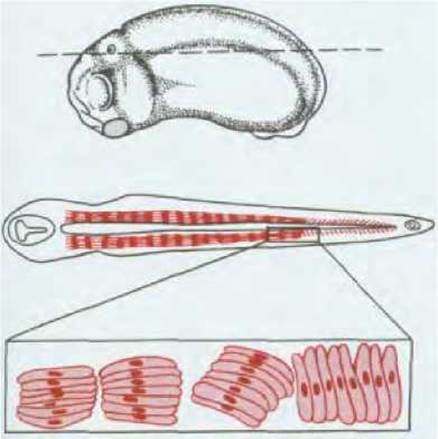
Рис. 16-16. Образование сомитов у Xenopus. Вверху - внешний вид зародыша при наблюдении сбоку. Прерывистой линией показана плоскость горизонтального разреза, изображение которого представлено в середине. Внизу схема перегруппировки клеток мезодермы в процессе формирования сомитов (при большом увеличении). У Xenopus все клетки, из которых потом образуются сомиты, исходно ориентированы под прямым углом к оси тела. В процессе формирования сомитов все клетки каждой группы поворачиваются одновременно.
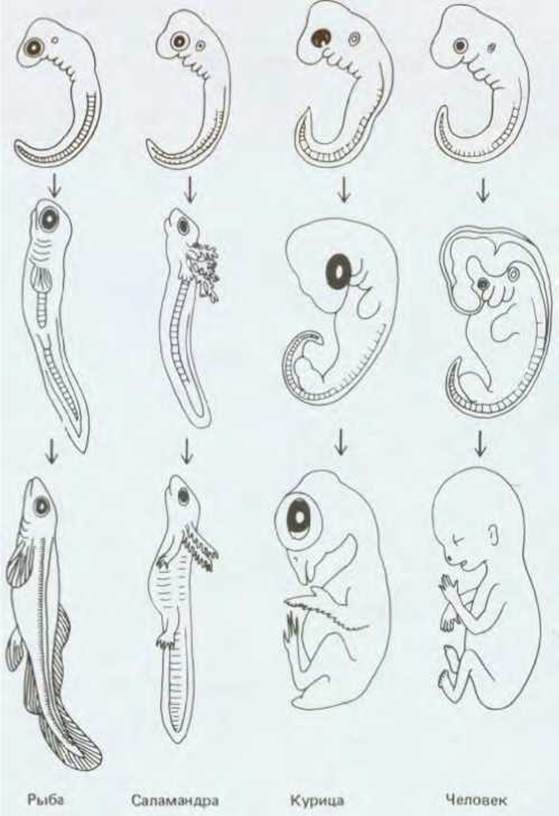
Рис. 16-17. Сравнение эмбриогенеза рыб, амфибий, птиц и млекопитающих. Ранние стадии развития (вверху) очень сходны; поздние стадии (внизу) различаются значительно сильнее. Самые ранние стадии изображены примерно в одинаковом масштабе, а более поздние стадии - в разном. (Е. Haeckel, Anthropogenic, oder Entwickelungsgechichte des Menschen. Leipzig, Engelmann, 1874.)
Сомиты обособляются не все сразу, а последовательно один за другим по направлению от головы к хвосту. Сегментация сопровождается изменением взаимных связей между клетками мезодермы. В ранней, несегментированной мезодерме сохраняется электрическое сопряжение клеток через щелевые контакты, исчезающие непосредственно перед или во время формирования сомитов. Очевидно, сначала изменяется характер взаимосвязи между клетками, затем они собираются в компактные группы и образуют сомиты (см. рис. 16-16). Позже мы увидим, что физическая сегментация коррелирует с появлением химических различий между соседними группами клеток сомитов (см. разд. 16.5.20), сегментирование которых напоминает расположение полосок у зебры. По всей вероятности, селективное сцепление, основанное на различиях в химизме поверхности клеток, является причиной разделения массива клеток на физически обособленные сомиты.
16.1.12. План строения тела позвоночного животного складывается в миниатюре на ранней стадии и сохраняется в период роста эмбриона [9]
Длина эмбриона на стадии формирования сомитов не превышает нескольких миллиметров, он обычно состоит из 105 клеток. До сих пор мы имели в виду Xenopus, но форма и размеры эмбриона примерно одинаковы и у саламандры, и у рыбы, и у курицы и у человека (рис. 16-17). На более поздних стадиях эмбрионы будут значительно различаться по величине и форме, но на этой стадии хорошо видно, что план строения у них один и тот же. Детали будут добавляться позже по мере роста зародыша. А пока центральная нервная система представлена нервной трубкой с утолщенным концом, из которого впоследствии разовьется головной мозг; кишечник и его производные заложены в виде энтодермальной трубки, сегментам туловища соответствуют сомиты. Некоторые типы соединительной ткани сердечно-сосудистой системы представлены более периферической несегментированной мезодермой, а эпидермальный слой кожи - эктодермой. В ходе последующего развития линейные размеры всех этих компонентов могут увеличиваться в 10, 100 и более раз, а суммарный объем и число клеток - в миллионы раз, но общий план строения тела остается прежним.
Заключение
Яйца большинства видов животных представляют собой довольно крупные клетки, содержащие запасы питательных веществ и других клеточных компонентов, синтез которых регулируется материнским геномом. У амфибий первым и весьма важным движением после оплодотворения является поворот кортекса яйца по отношению к его внутреннему содержимому. Асимметрия, создаваемая этим поворотом, равно как и исходная асимметрия в распределении содержимого в яйце перед оплодотворением, определяет будущие передне-заднюю и дорсовентральную оси тела. В ходе последующих делений дробления яйцо делится на множество более мелких клеток, однако общий объем зародыша не увеличивается. Клетки наружных слоев эмбриона формируют между собой плотные контакты, изолируя внутреннее содержимое эмбриона от окружающей среды. Внутрь эмбриона проникает жидкость, в результате чего в нем формируется полость (бластоцель). Стенка такого зародыша называется бластодермой, а сам зародыш носит название бластулы.
В процессе гаструляции бластодерма на одном конце зародыша начинает втягиваться внутрь вследствие изменения формы ее клеток. Полагают, что движущей силой последних этапов гаструляции является конвергентное расширение впячиваемого эпителия за счет изменения упаковки его клеток, что трансформирует эмбрион в трехслойную структуру с внутренней трубкой (энтодермой), наружным покровом (эктодермой) и промежуточным слоем клеток, который отделился от этой эктодермы (мезодермой). Из энтодермы в дальнейшем сформируется выстилка кишки и ее производных, из эктодермы - главным образом эпидермис и нервная система, а из мезодермы - большая часть мышц и соединительной ткани, сердечно-сосудистая система и мочеполовой тракт. В результате гаструляции сближаются и взаимодействуют между собой группы клеток, находившиеся ранее вдали друг от друга. Например, дорсальная мезодерма вызывает утолщение эктодермы, теперь лежащей над нею; данный участок сворачивается и отделяется, формируя нервную трубку и нервный гребень. Этот процесс, именуемый нейруляцией, также зависит от изменения формы эктодермальных клеток. В средней части дорсальной мезодермы расположен тяж специализированных клеток, именуемый хордой; этот тяж образует центральную ось эмбриона. Удлиненные массы мезодермы, лежащие по бокам от хорды, фрагментируются и дают начало сомитам, из которых возникают позвонки и скелетная мускулатура.