Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Основы клеточной теории
Митохондрии и пластиды
Митохондрии, которые имеются у всех клеток эукариот как автотрофных (у фотосинтезирующих растений), так и гетеротрофных (у животных и грибов), содержат ферменты и другие белково-мембранные структуры для окисления органических соединений и синтеза аденозин-трифосфата (АТФ) - главного источника энергии внутриклеточных биохимических превращений.
Митохондрии являются одними из самых больших клеточных органелл с размерами порядка размеров бактерии Е. coli. В некоторых эукариотических клетках митохондрии могут занимать до 25% цитозоля.
Количество митохондрий в клетках значительно варьируется в зависимости от потребностей в энергии данного типа клеток. В клетках некоторых типов водорослей и простейших содержится лишь одна митохондрия, в сперматозоидах различных видов животных - 20-70, а в клетках тела млекопитающих - от 100 до нескольких тысяч. Больше всего митохондрий содержится в клетках тканей, постоянно использующих много энергии для выполнения своих функций - в мышцах, в печени и в почках. Митохондрии могут занимать от 2 до 48% от общего объема клетки, но в среднем эта величина составляет 18%.
Митохондрия сформирована двумя структурно и функционально различными мембранами - внешней и внутренней, разделенными межмембранным пространством (рисунок 20).

Рисунок 20 - Схема митохондрии
Внутренняя мембрана, на которой сосредоточены ферменты, катализирующие окислительные реакции в дыхательной цепи и синтезирующие АТФ, образует многочисленные складки-гребни (кристы - cristae), которые вогнуты внутрь матрикса - центральной части митохондрии.
Митохондрия содержит свои собственные ДНК и рибосомы, но большинство белков она получает из цитоплазмы клетки. Чтобы подчеркнуть роль митохондрий, как источника АТФ, их часто называют "энергетическими станциями" (powerhouses) клетки.
Наружная мембрана митохондрии содержит множество белков-поринов, которые пропускают в межмембранное пространство ионы и малые молекулы-метаболиты (с массой до 5000 Да).
Главным барьером на пути метаболитов в матрикс митохондрии является внутренняя мембрана митохондрии. Внутренняя мембрана содержит множество мембранных белков (до 75% массы внутренней мембраны), одни из которых осуществляют необходимые биохимические преобразования, а другие (транспортёры) переносят метаболиты через мембрану.
Во внутренней мембране повышенная концентрация кардиолипина (дифосфатидил глицерол), за счет чего снижена её проницаемость для протонов, и на внутренней мембране создается значительная протондвижущая сила.
Новые митохондрии образуются только делением или почкованием имеющихся митохондрий.
Пластиды встречаются только у фотосинтезирующих эукариотических организмов (высшие растения, водоросли, некоторые одноклеточные простейшие). По пигментному составу, связанному с их функциями, различают три основных типа пластид: хлоропласты (зеленые пластиды), хромопласты (желтые или оранжевые пластиды) и лейкопласты (бесцветные пластиды). В каждой клетке встречаются пластиды только одного типа.
Хлоропласты содержат зеленый пигмент хлорофилл и осуществляют на свету фотосинтез, в результате которого энергия света превращается в химическую энергию и используется для синтеза нужных растению веществ. Хлоропласты встречаются почти во всех клетках растения, куда проникает свет, но особенно много их в клетках листьев.
Лейкопласты встречаются в тех клетках растения, куда не проникает солнечный свет. Их главная функция - накопление и хранение запасных питательных веществ.
Хромопласты накапливают желтые, оранжевые и красные пигменты (каротиноиды). Хромопласты так же, как и окрашенные вакуоли, придают яркий цвет некоторым частям растений. Например, хромопласты придают цвет лепесткам лютика, одуванчика или тюльпана, зрелым плодам томатов, рябины, арбуза или апельсина, моркови и свекле.
Пластиды способны превращаться друг в друга. Если освещать бесцветную подземную часть растения, то лейкопласты под действием света начнут накапливать хлорофилл и превратятся в хлоропласты.
По этой причине, например, зеленеет оставленный на солнце клубень картофеля. Хлоропласты же, старея, теряют свою зеленую окраску и превращаются в хромопласты. Такой процесс происходит в листьях осенью: зеленый хлорофилл в хлоропластах начинает разрушаться и тогда становятся видны каротиноиды, которые всё время присутствовали в хлоропластах, но были замаскированы хлорофиллом. В результате зеленые листья становятся желтыми, оранжевыми или красными. Взаимные превращения пластид позволяют рассматривать их как различные типы дифференцировки одного типа органеллы. Это подтверждается общностью строения разных пластид.
Основным типом пластид являются хлоропласты (рисунок 21). Они обычно имеют постоянную форму вытянутого эллипсоида вращения с длинной и короткой полуосями 2-3 и 0,5-1,5 мкм соответственно.
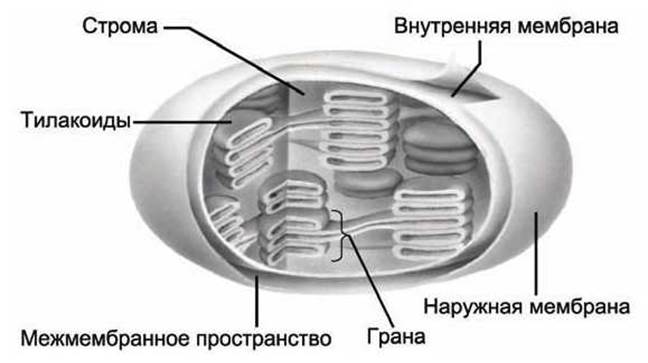
Рисунок 21 - Схема хлоропласта
Большинство растений имеют хлоропласты объемом от 30 мкм3 до 60 мкм3. (Мелкие пластиды (меньше 30 мкм3) найдены у тополя, осины, ясеня, некоторых видов осоки. Очень крупные хлоропласты типичны для тенелюбивых растений, например у копытня - 227 мкм3, заячьей капусты - 240 мкм3, фиалки - 100 мкм3).
Число хлоропластов на клетку варьирует от 3 до 400, обычно в среднем на клетку приходится 10-30 хлоропластов. (Более 400 пластид на
клетку найдено у табака, ревеня, у некоторых видов лилий. Малопластидными являются, например, клетки тростника (4-10), земляники (8-12), лимона (5-10)). Число пластид в клетке в значительной мере коррелирует с содержанием ДНК в ядре и гораздо слабее с объемом клетки. Для оценочных расчетов можно использовать среднестатистическую величину - 14 хлоропластов на 10 тыс. мкм3 объема клетки.
Хлоропласты, так же как и митохондрии, имеют две мембраны: внешнюю, которая содержит порины и проницаема для метаболитов, и внутреннюю, которая содержит транспортные белки, обеспечивающие обмен метаболитами с цитозолем клетки. Обе эти мембраны хлоропластов в фотосинтезе не участвуют.
В отличие от митохондрий, хлоропласты имеют ещё одну, третью мембрану - мембрану тилакоидов.
В зрелом хлоропласте высших растений различают два типа внутренних мембран: (1) плоские протяженные складки, образованные внутренней мембраной оболочки и способные простираться почти через всю пластиду, - ламеллы стромы и (2) плоские замкнутые мембранные мешки, имеющие форму диска, - тилакоиды.
Часто тилакоиды образуют стопки наподобие столбика монет, называемые гранами. Число тилакоидов на одну грану может варьировать от нескольких штук до 50 и более.
Количество гран в хлоропластах может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются. В состав граны, кроме тилакоидов, обычно входят и участки ламелл стромы, которые таким образом связывают между собой отдельные граны хлоропласта.
Все тилакоиды хлоропласта, собранные в граны имеют единую тилакоидную мембрану, которая разделяет тилакоидный люмен - внутренний объем тилакоидов, представляющий единый компартмент, - и строму - другой компартмент хлоропласта между тилакоидами и внутренней мембраной хлоропласта.
В тилакоидной мембране расположены множество интегральных мембранных белков, содержащих специфические простетические группы и светопоглощающие пигменты, например, хлорофилл.
Синтез углеводов происходит в строме хлоропласта. В фотосинтетических бактериях складки плазматической мембраны формируют мембранные структуры, в которых локализованы системы фотосинтеза и которые также называют тилакоидными мембранами.
В строме хлоропластов обнаруживаются молекулы пластидной ДНК, рибосомы и различные включения. Среди включений наиболее распространенными являются крахмальные зерна и пластоглобулы.
Отложение крахмальных зерен тесно связано с функционированием хлоропластов: часть образующихся в процессе фотосинтеза углеводов идет на синтез крахмала в строме. Крахмальные зерна имеют овальную, сферическую или неправильную форму, характерную для каждого вида растения, имеют плотность 1,5—1,6 г/см3 и обладают двойным лучепреломлением. Диаметр крахмальных зерен варьирует от 0,2 до 7 мкм, когда крахмальные зерна занимают почти весь объем пластиды, обычно лейкопласта, называемого в этом случае амилопластом. (Самые крупные крахмальные зерна у картофеля, а самые мелкие - у риса и гречихи. Размер крахмальных зерен, как правило, непостоянен).
Пластоглобулы имеют диаметр от 0,2 до 1 мкм в зависимости от вида растения. Их число и величина в общем случае увеличиваются с возрастом хлоропластов, достигая максимума в период осеннего отмирания листьев.
Быстрая сборка мембран тилакоидов при преобразовании лейкопластов в хлоропласты на свету обычно сопровождается уменьшением числа пластоглобул. В этой связи пластоглобулы рассматриваются как резервный фонд липидов пластидных мембран.
Строение хлоропластов низших фотосинтезирующих растений (зеленые, бурые и красные водоросли), называемых хроматофорами, в общих чертах сходно со строением хлоропластов клеток высших растений.
Водоросли часто содержат по одному большому хроматофору на клетку, форма которого может быть очень разнообразна, например, длинная спиральная лента или разветвленная сеть. Так же, как у высших растений, хроматофоры окружены оболочкой из внешней и внутренней мембран, последняя из которых образует внутренние впячивания в виде параллельных плоских мешков.
Однако граны в хроматофорах водорослей не встречаются. У зеленых водорослей в состав хроматофора входят пиреноиды, представляющие собой окруженные мелкими пузырьками зоны, вокруг которых происходит отложение крахмала.
Так же, как новые митохондрии, новые пластиды в клетке образуются только за счет деления или почкования уже существующих. Увеличение числа пластид обычно происходит за счет деления не имеющих развитой структуры пропластид или инициальных частиц с последующим развитием и дифференцировкой их в хлоропласта или лейкопласты.
Однако в молодых растущих клетках делятся не только пропластиды, но и хлоропласта с хорошо развитой внутренней мембранной системой.
При созревании хлоропластов, когда в них накапливается большая масса пластоглобул, их способность к делению утрачивается. Тем не менее обычно клетка на протяжении всей её жизни поддерживает фонд хлоропластов, имеющих более мелкие размеры и сохраняющих способность делиться. В молодых листьях такие хлоропласта составляют 5%, у зрелых - 1% от общего числа пластид.
Как правило, деление и рост клетки сопровождаются делением хлоропластов. Деление хлоропластов прекращается, когда клетка достигает максимальных размеров. В то же время рост клетки и деление хлоропластов относительно независимы друг от друга.
В клетках прокариот нет внутренних мембран, но плазматическая мембрана фотосинтетических бактерий содержит мезосомы - внутрицитоплазматические мембранные структуры везикулярной и трубчатой формы, образующиеся путём впячивания плазматической мембраны внутрь цитоплазмы.