Основы молекулярной биологии. Часть 1: Молекулярная биология клетки - А.Н. Огурцов 2011
Окисление глюкозы и жирных кислот
Аэробное окисление в митохондриях
Митохондрии являются одними из самых больших клеточных органелл с размерами порядка размеров бактерии Е. coli. В некоторых эукариотических клетках митохондрии могут занимать до 25% цитозоля.
Митохондрия имеет две мембраны: проницаемую для метаболитов внешнюю мембрану и внутреннюю мембрану, на которой расположены системы переноса электронов и синтеза АТФ (рисунок 20).
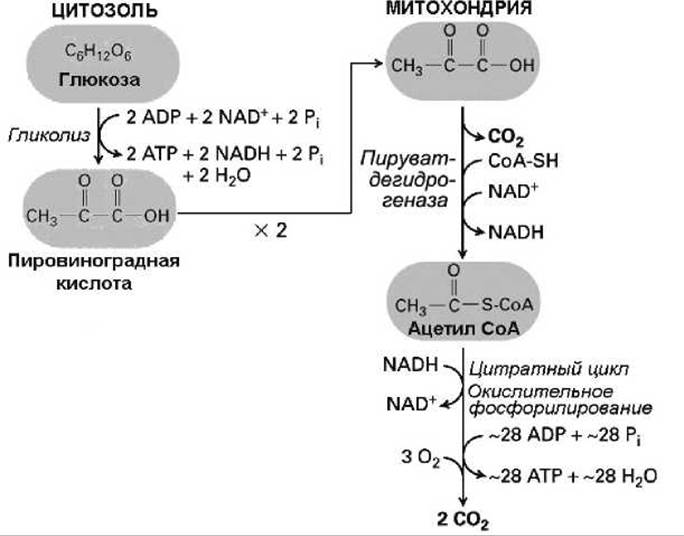
Рисунок 153 - Схема аэробного метаболизма
Большинство реакций окисления пирувата и жирных кислот до СО2 и Н2О и сопряженного синтеза АТФ из АДФ и Рi проходят на внутренней мембране митохондрии, в её кристах и матриксе (рисунок 154).
Реакции окисления пирувата и жирных кислот до СO2 и Н2O и сопряженного синтеза АТФ состоят из множества этапов, но они могут быть разделены на три группы реакций, каждая из которых происходит в определенном месте на внутренней мембране митохондрии:
1. Окисление пирувата и жирных кислот до СO2 сопряженное с восстановлением (а) НАД⊕ до НАДН (рисунок 151), и (б) ФАД (флавинадениндинуклеотид) до ФАДН2 (рисунок 155). Эти переносчики электронов часто называют коферментами. Большинство реакций этой группы происходят в матриксе, две из них катализируются мембранными белками внутренней мембраны, каталитические центры которых ориентированы внутрь матрикса.
2. Электроны переносятся от НАДН и ФАДН2 к O2, регенерируя НАД⊕ и ФАД. Эти реакции происходят на внутренней мембране и приводят к созданию протондвижущей силы.
3. Протонный градиент используется для синтеза АТФ комплексом F0F1 (АТФ-синтаза) на внутренней мембране.

Рисунок 154 - Схема аэробного окисления пирувата и жирных кислот в митохондрии: 1 - пируват дегидрогеназа, нитратный цикл и метаболизм жирных кислот; 2 - транспорт электронов от НАДН и ФАДH2 к O2 и создание протондвижущей силы; 3 - синтез АТФ АТФ-синтазой (которую иногда называют комплекс V)
Кристы значительно увеличивают площадь внутренней мембраны митохондрии, соответственно увеличивая выход АТФ. В типичной митохондрии клетки печени площадь внутренней мембраны в пять раз больше площади наружной мембраны. А в митохондриях сердечной мышцы или скелетных мышц (в которых происходит повышенный расход АТФ) площадь внутренней мембраны ещё в три раза больше, чем в клетках печени.
В растениях углеводы хранятся в виде крахмала, который гидролизуется до глюкозы. Затем в результате гликолиза синтезируется пируват, молекулы которого так же, как и в клетках животных, переносятся в митохондрии. Митохондриальное окисление пирувата, сопровождающееся синтезом АТФ, происходит и в фотосинтезирующих клетках, когда отсутствует освещение (темновые реакции), а в корнях и других нефотосинтезирующих тканях - всё время.

Рисунок 155 - Схема окисления флавинадениндинуклеотида: а - окисленная форма ФАД; б - восстановленная форма ФАДH2. Рибит: -СH2(HСОН)3СH2-
После транспортировки пирувата в матрикс фермент пируват дегидрогеназа (используя кофермент СоА) катализирует его превращение в ацетил-СоА (рисунок 156).
В этой реакции, в отличие от реакций, катализируемых лактатдегидрогеназой и алкогольдегидрогеназой (рисунок 152), НАД⊕ восстанавливается до НАДН.