Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Синтез белков на рибосомах
Инициация трансляции
Подобно транскрипции, сложный процесс трансляции может быть разделен на три стадии - инициация, элонгация и терминация. Рассмотрим трансляцию в клетках эукариот, имея ввиду, что механизм трансляции в общих чертах одинаков во всех организмах.
В большинстве мРНК стартовым является AUG - кодон метионина. Поэтому распознавание этого кодона является критически важным для установления правильной рамки считывания (reading frame) для всей мРНК.
И в прокариотах, и в эукариотах существует две различных метиониновых тРНК:
1) tRNAiMet - может инициировать синтез белка,
2) tRNAMet - способна только добавлять метионин в уже растущую полипептидную цепь.
Одна и та же аминоацил-тРНК-синтетаза (MetSR) аминоацилирует обе тРНК метионином. Но только Met-tRNAiMet (т.е. tRNAiMet с присоединенным активированным метионином) может связаться с особым активным центром, Y-центром, малой рибосомной субъединицы, чтобы начать синтез полипептидной цепи. Обычная Met-tRNAMet, равно как и остальные аминоацилированные тРНК, связывается только с другим активным центром рибосомы, А-центром.
После терминации предыдущей трансляции диссоциировавшие 40S и 60S субъединицы рибосомы связываются с факторами инициации eIF3 и eIF6, соответственно, что делает их готовыми к следующему циклу трансляции (рисунок 41).
В обозначении "eIF" символ "е" означает эукариотический фактор, "I" означает процесс инициации, a "F" - сокращение слова "фактор".
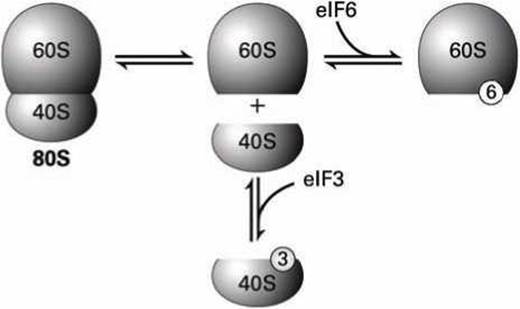
Рисунок 41 - Связывание субъединиц рибосомы с факторами инициации
В ходе первого этапа трансляции происходит сборка комплекса из рибосомной субъединицы, мРНК и активированной инициирующей тРНК, которая правильно позиционируется на стартовом кодоне.
Прединициаторный комплекс трансляции формируется, когда комплекс (40S-субъединица-еIРЗ) связывается с eIF1A и тройным комплексом (Met-tRNAiMet-eIF2-GTP) (рисунок 42, этап 1) (GTP - гуанозин трифосфат).
Клетка может регулировать белковый синтез с помощью фосфорилирования серина в факторе инициации eIF2, связанном с GTP, в результате чего фосфорилированный eIF2 не способен заменить уже использованный GDP на GTP и, следовательно, не может связаться с Met-tRNAiMet. Это ингибирует синтез белка.
В ходе инициации трансляции 5'-кэп мРНК связывается eIF4E субъединицей кэп-комплекса eIF4.
Комплекс (MPHK-eIF4) затем объединяется с прединициаторным комплексом, благодаря взаимодействию еIF4G-субъединицы и eIF3, образуя инициаторный комплекс (рисунок 42, этап 2).

Рисунок 42 - Этапы инициации трансляции
Затем инициаторный комплекс сдвигается вдоль - сканирует - мРНК, при этом субъединица eIF4A, обладающая геликазной активностью, расплетает вторичную структуру мРНК, используя энергию гидролиза АТФ.
Сканирование прекращается, когда антикодон Met-tRNAiMet распознает стартовый кодон, которым является первый AUG вниз от 5'-конца в большинстве эукариотических мРНК (рисунок 42, этап 3).
Распознавание стартового кодона приводит к гидролизу GTP, связанному с eIF2. Это необратимое событие исключает дальнейшее сканирование мРНК.
Выбор инициирующего кодона AUG облегчается его специфическим нуклеотидным окружением, которое называется последовательностью Козак ("Kozak sequence", в честь Марилин Козак, которая открыла её):
![]()
Первый А и последний G в этой последовательности оказывают самое сильное влияние на эффективность инициации трансляции.
Сразу после того, как малая рибосомная субъединица с присоединенным Met-tRNAiMet правильно расположится на стартовом кодоне, присоединение к ней большой (60S) субъединицы завершает формирование 80S-рибосомы.
Это требует участие ещё одного фактора eIF5 и гидролиза связанного с ним GTP (рисунок 42, этап 4).
То, что реакция объединения большой и малой субъединиц связана с гидролизом GTP, делает её необратимым шагом, и поэтому рибосомные субъединицы не диссоциируют до тех пор, пока вся мРНК не транслируется и не произойдет терминация синтеза белка.
Во время элонгации растущий полипептид остаётся присоединенным к P-центру рибосомы.
В большинстве эукариот описанный выше синтез протеинов начинается на участке порядка 100 нуклеотидов от кэпированного 5'-конца мРНК. Однако в некоторых клеточных мРНК существует так называемый внутренний участок связывания рибосомы, IRES (internal ribosome entry site), расположенный далеко от 5'-конца. Кроме того, трансляция некоторых вирусных мРНК, в которых отсутствует 5'-кэп, инициируется на IRES не изученными пока механизмами взаимодействия инфицированной клетки с вирусом.
По-видимому, те же факторы инициации трансляции, которые способствуют сканированию рибосомы от 5'-конца в поисках AUG, необходимы и для определения AUG во внутреннем участке связывания рибосомы, но деталей идентификации IRES пока не известно.
Предварительно есть основания предполагать, что IRES свернут в особую структуру, которая присоединяется к третьему активному центру рибосомы, Е-центру, тем самым рибосома правильно устанавливается центром Р напротив внутреннего AUG стартового кодона.