Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Репликация и репарация ДНК
Модели репликации
После расшифровки структуры двойной спирали ДНК Уотсоном и Криком было предположено, что синтез новых спиралей происходит с использованием уже существующих (родительских) спиралей в качестве матрицы и формировании дочерних спиралей, комплементарных родительским.
Такая модель матричного синтеза пар нуклеотидов теоретически может быть реализована либо по консервативному, либо по полуконсервативному механизму.
Согласно консервативному механизму, две дочерние нити могут образовать новую двойную спираль ДНК, или дуплекс (duplex), при этом родительский дуплекс будет сохранен.
Согласно полуконсервативному механизму, цепи спирали родительской ДНК полностью разъединяются, каждая из них образует новый дуплекс с дочерними цепями, которые образуются на родительских цепях путем спаривания комплементарных нуклеотидов.
Доказательство того, что ДНК реплицируется по полуконсервативному механизму, было получено в классическом эксперименте представленном на рисунке 50, который провели Мэтт Мезельсон (Mathew Meselson) и Фрэнк Сталь (Franklin Stahl) в 1958 году.
Исходные клетки Е. coli были выращены в среде содержащей только "тяжелый" изотоп азота 15N, поэтому все исходные ДНК были "мечены" этим изотопом.
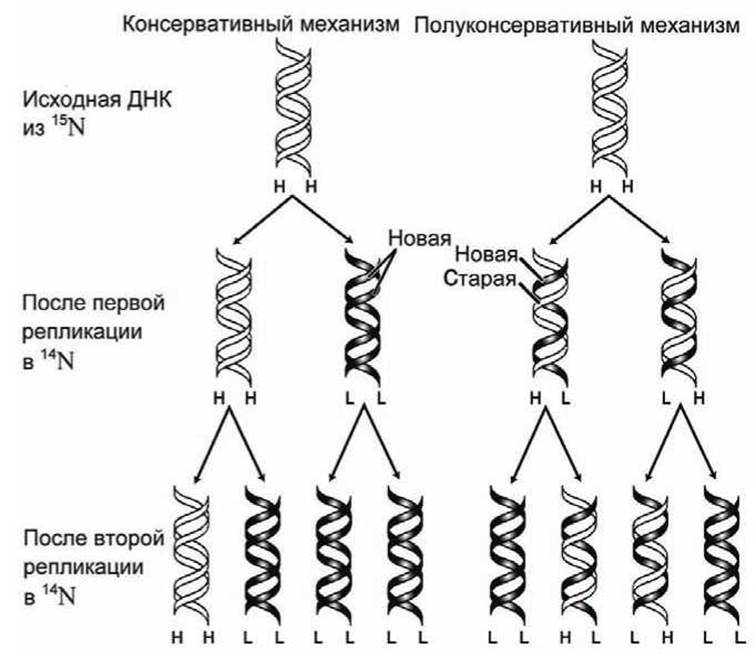
Рисунок 50 - Схемы консервативного и полуконсервативного механизмов репликации ДНК
Затем, клетки были перенесены в среду, содержащую только "легкий" изотоп азота 14N. Периодически отбирались образцы, и ДНК каждого образца анализировалась методом центрифугирования в равновесном градиенте плотности. Это давало возможность зарегистрировать тяжелые (heavy-heavy, Н-Н), смешанные (heavy-light, Н-L) и легкие (light-light, L-L) дуплексы.
Если бы репликация проходила по консервативному механизму, то невозможно было бы образование Н-L дуплексов, в то время как полуконсервативный механизм в большинстве генерирует как раз H-L дуплексы (рисунок 50). Эксперимент показал, что первое дочернее поколение состоит из Н-L дуплексов, а во втором (точнее, в 1,9-м) примерно поровну Н-L и L-L дуплексов (рисунок 51). В дальнейших поколениях появлялась все большая доля L-L дуплексов, как и предсказывал полуконсервативный механизм репликации.

Рисунок 51 - Анализ репликации ДНК методом центрифугирования
Таким образом, копирование матричной цепи в комплементарную цепь является универсальной особенностью и репликации ДНК и транскрипции ДНК в РНК. В обоих случаях информация в матричной цепи сохраняется.
В некоторых вирусах одноцепочечные молекулы РНК выполняют функцию матрицы для синтеза комплементарных ДНК или РНК цепей. Однако подавляющее большинство молекул РНК и ДНК в клетках синтезируются из имеющихся дуплексов.
Подобно РНК, молекулы ДНК синтезируются из дезоксинуклеозид- 5'-трифосфатов (дНТФ). Аналогично синтезу РНК, синтез ДНК идет в направлении 5'→3', поскольку, как и при синтезе РНК (рисунок 8), рост цепи ДНК идет за счет формирования фосфодиэфирной связи между З'-кислородом растущей цепи и а-фосфатом дНТФ .
Синтез нити ДНК катализируется ферментом ДНК-полимераза. Субстратами ДНК-полимеразы являются четыре дезоксирибонуклеозид трифосфата: дАТФ, дЦТФ, дГТФ, и дТТФ.
Как было сказано ранее, РНК-полимераза сама находит соответствующую стартовую точку на дуплексе ДНК и инициирует синтез нити РНК, комплементарной матричной ДНК-нити.
В отличие от РНК-полимеразы, ДНК-полимераза не может самостоятельно инициировать новый (de novo) синтез цепи. Для этого ей нужно, чтобы уже существовала цепь или ДНК, или РНК, которая называется праймер (рисунок 52).
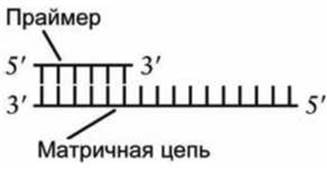
Рисунок 52 - Схема праймера репликации
Когда уже есть праймер, спаренный с матричной нитью ДНК, тогда ДНК-полимераза катализирует реакцию присоединения того дНТФ к гидроксильной группе на 3'-конце праймера, который закодирован в матричной цепи.
Если праймером служит РНК, то дочерняя цепь является РНК на 5'-конце и ДНК на 3'-конце.