Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Репликация и репарация ДНК
Двунаправленная репликация
Как показано на рисунках 53 и 54, обе родительские нити после расплетания и разъединения репликационной вилкой копируются в дочерние нити.
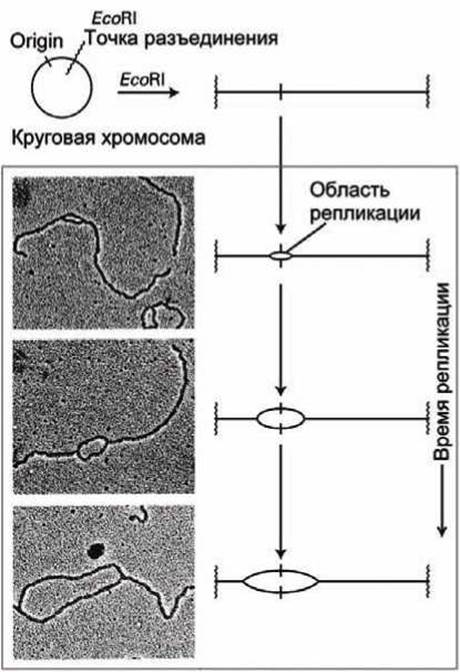
Рисунок 55 - Двунаправленная репликация ДНК
С одного ориджина может начинаться как единственная репликация, при которой репликационная вилка перемещается в одном направлении вдоль родительской спирали, так и двунаправленная репликация (bidirectional growth) (рисунок 55), при которой две репликационные вилки организуются на одном и том же ориджине и двигаются в противоположных направлениях. Двунаправленный рост наблюдался во многих экспериментах, включая представленный на рисунке 55.
Кольцевая ДНК вируса была разрезана рестрикционным ферментом ЕсоRI, и затем наблюдался рост области репликации симметрично в обе стороны от ориджина, демонстрируя двунаправленную репликацию.
В настоящее время считается, что все прокариотические и эукариотические клетки задействуют процесс двунаправленной репликации.
Так, в рассмотренном ранее примере, двунаправленная репликация SV40 ДНК инициируется связыванием двух больших Т-антигеновых геликаз с одним SV40 ориджином и присоединением остальных необходимых белков так, что формируются две репликационные вилки.
Затем эти две вилки перемещаются в противоположных направлениях, начиная от ориджина. Как показано на рис 56, левая репликационная вилка осуществляет синтез ДНК справа налево, а правая - слева направо.
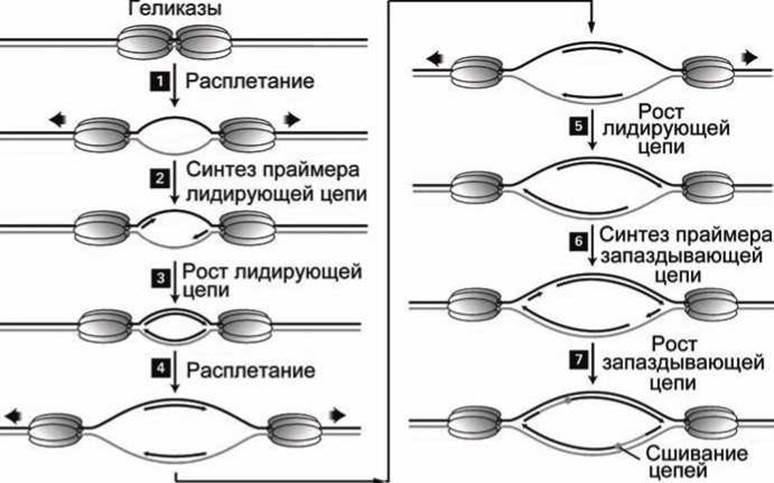
Рисунок 56 - Схема двунаправленной репликации ДНК
Используя энергию гидролиза АТФ, геликазы двигаются в противоположных направлениях, образуя две однонитевые цепи, стабилизированные RPA белками (рисунок 56, этап 1).
Комплексы (праймаза-Pol а) синтезируют короткие праймеры на каждой из родительских матричных цепей (рисунок 56, этап 2).
Комплексы (PNCA-Rfc-Pol δ) замещают комплексы (праймаза-Роl а) и наращивают короткие праймеры, образуя лидирующую цепь на каждой из репликационных вилок (рисунок 56, этап 3).
Геликазы продолжают расплетать дуплекс, и RPA белки связываются с новыми свежерасплетёнными однонитевыми участками (рисунок 56, этап 4).
Комплексы (PCNA-Rfc-Pol δ) продолжают наращивать лидирующие цепи (рисунок 56, этап 5).
Комплексы (праймаза-Pol а) синтезируют праймеры для синтеза запаздывающих цепей на обеих репликационных вилках (рисунок 56, этап 6).
Комплексы (PCNA-Rfc-Pol δ) замещают комплексы (праймаза-Роl а) и наращивают фрагменты Оказаки запаздывающих цепей, которые в конце объединяются с 5'-концами лидирующих цепей. Место сшивки показано кружком (рисунок 56, этап 7).
Репликация продолжается дальнейшим повторением этапов 4-7, т.е. расплетанием ДНК и синтезом лидирующих и запаздывающих цепей.
Поскольку при репликации область ориджина копируется в дочерние цепи в первую очередь, то возможна инициация нового раунда репликации ещё до завершения предыдущего, и возникает разветвленная структура, показанная на рисунке 57.
Такая "дихотомическая" репликация позволяет бактериям при благоприятных условиях иметь время генерации генома меньшее, чем время, необходимое на завершение раунда репликации ДНК.
В отличие от SV40 ДНК, эукариотические хромосомные молекулы ДНК содержат множество ориджинов репликации, разделенных сотнями и тысячами килобэйзов (kb, kilobase, тысяча пар оснований). Шестисубъединичный белок, который называется комплекс распознавания ориджина, ORC (origin recognition complex), связывается с каждым ориджином и присоединяет к себе белки, необходимые для того, чтобы связать с ДНК клеточные гексамерные геликазы, состоящие из шести гомологичных МСМ-белков.
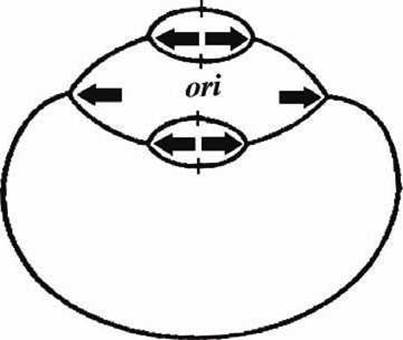
Рисунок 57 - Дихотомическая репликация бактериальной хромосомы
Две противоположно направленные MCM-геликазы расплетают родительский дуплекс в ориджине. Белки RPA (replication protein А), связываясь с разъединенными одиночными нитями, оптимально ориентируют их для синтеза дочерних цепей. Синтез праймеров и последующие шаги в репликации клеточной ДНК происходит, как полагают в настоящее время, таким же образом, как и при репликации SV40 ДНК (рисунки 53 и 54).
Репликация клеточной ДНК и другие события, приводящие к размножению клеток, имеют тонкие механизмы регуляции, так, чтобы необходимое количество клеток каждой ткани создавалось в процессе роста и всей жизни организма. Так же, как и при транскрипции большинства генов, контроль инициации является главным механизмом регуляции репликации ДНК клеток. Активация МСМ-геликаз, которые необходимы для инициации репликации ДНК, регулируется специфическими протеин-киназами, которые называются циклин-зависимые киназы S-фазы (S-phase cyclin-dependent kinases). Другие циклин-зависимые киназы регулируют дополнительные аспекты клеточного деления, включая сложный процесс митоза, с помощью которого эукариотические клетки делятся на две дочерние клетки.