Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Клонирование ДНК
Технология рекомбинантных ДНК
Для того, чтобы исследовать структуру и функции гена на молекулярном уровне необходимо уметь получать большое количество определённого гена в чистой форме. Существует много методов под общим названием "Технология рекомбинантных ДНК", которые используются для клонирования ДНК - получения большого числа идентичных копий исходного гена.
Рекомбинантной ДНК называется молекула ДНК составленная из участков ДНК, полученных из разных источников.
Основной задачей при клонировании интересующего нас фрагмента ДНК является включение его в состав так называемого вектора.
Вектором называется фрагмент молекулы ДНК, который имеет все необходимые компоненты для своей репликации в той клетке, которая будет использована для размножения.
После того, как рекомбинантная молекула ДНК, состоящая из вектора и внедренного фрагмента ДНК, который и должен быть клонирован, вводится в клетку, внедренный фрагмент ДНК реплицируется вместе с вектором, образуя в итоге большое число идентичных молекул ДНК. Общая схема клонирования представлена на рисунке 83.
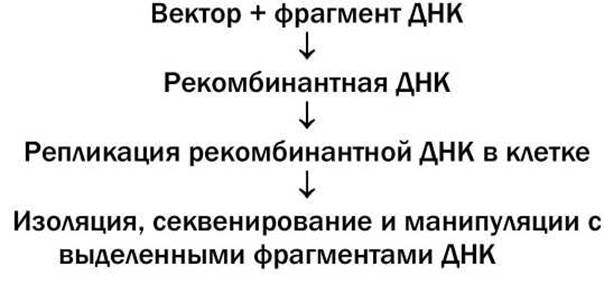
Рисунок 83 - Общая схема клонирования ДНК
Хотя изобретено множество экспериментальных приемов, варьирующих реализацию каждого из этапов, в целом клонирование ДНК содержит в себе все эти существенные этапы.
Ниже мы рассмотрим два типа векторов, которые наиболее часто используются в клетках Е. coli:
1) плазмидные векторы, которые реплицируются в клетке вместе с клеточным геномом,
2) векторы на основе бактериофага X, которые реплицируются как литические вирусы, убивая, в результате, клетку и упаковывая реплицированные ДНК в вирионы.
Главная задача при клонировании ДНК состоит в получении коротких участков ДНК организма, содержащих специфические гены. Кроме того, только относительно небольшие молекулы ДНК могут быть включены в состав существующих векторов. Поэтому очень длинные молекулы ДНК генома организма должны быть разъединены на фрагменты нужной длины, которые затем могут быть присоединены к ДНК- векторам.
Для синтеза таких рекомбинантных ДНК используют два типа ферментов
1) рестрикционные ферменты,
2) ДНК-лигазы.
9.1.1. Фрагментирование ДНК. Рестрикционные ферменты - это эндонуклеазы, синтезируемые бактериями, которые распознают специфические участки ДНК из 4-8 пар нуклеотидов, называемые участком рестрикции или сайтом рестрикции (restriction site), и разрезают обе нити ДНК в этом месте.
Сайт рестрикции обычно представляет собой палиндромную последовательность (palindromic sequence) - участок двухцепочечной молекулы ДНК, обе цепи которого обладают одинаковой последовательностью нуклеотидов при прочитывании от 5'- к 3'-концу, т. е. палиндромная последовательность является тандемным инвертированным повтором, например:
![]()
Для каждого рестрикционного фермента бактерия синтезирует соответствующий модифицирующий фермент, который защищает собственную ДНК бактерии от разрезания с помощью модификации её в точке потенциального разрезания. Модифицирующий фермент добавляет метальную группу к одному или двум основаниям, как правило, в пределах сайта рестрикции. Когда такое метилирование ДНК проведено, рестрикционная эндонуклеаза не может разрезать ДНК.
Модифицирующие (метилирующие) ферменты вместе с рестрикционными эндонуклеазами образуют рестрикционно-модифицирующую систему, которая защищает собственную ДНК и разрушает чужеродную ДНК (например, ДНК бактериофагов или ДНК, присоединенную в ходе трансформации), разрезая её во всех сайтах рестрикции.
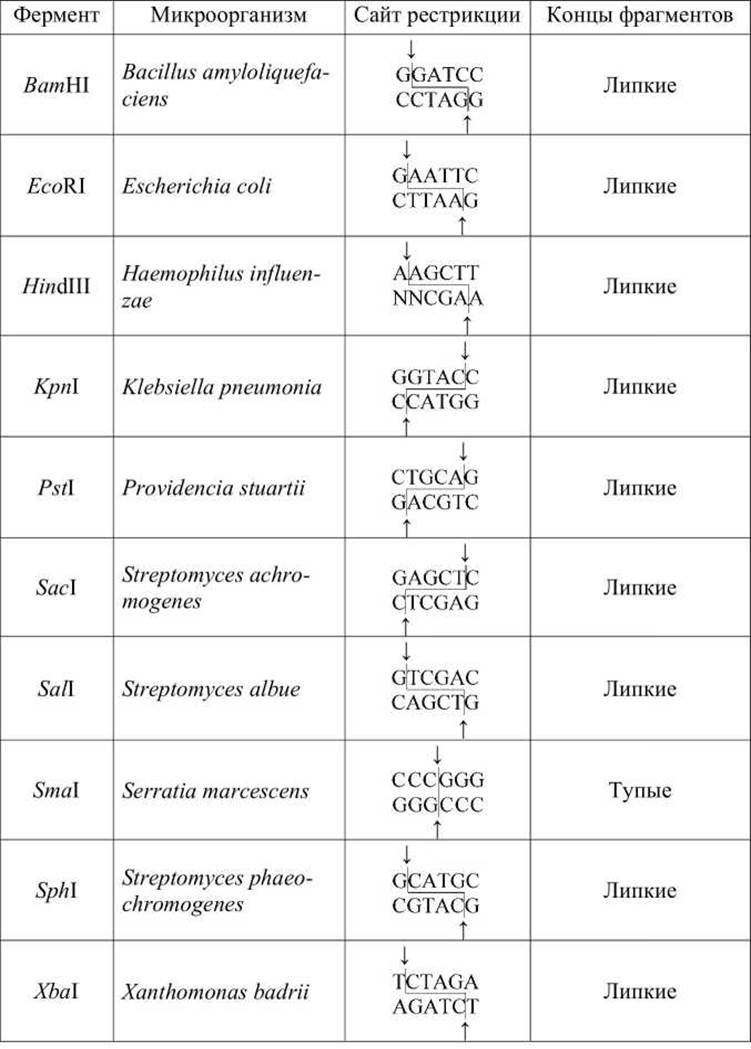
Большинство рестрикционных ферментов производят ступенчатый разрез (или разрез уступом, staggered cut) двух цепей ДНК в сайте рестрикции, образуя фрагменты, имеющие одноцепочечные "хвосты" на концах (рисунок 84, таблица 5).

Рисунок 84 - Разрезание ДНК рестрикционным ферментом EcoRI
Эти хвосты фрагментов, образующиеся при разрезании ДНК в данном сайте рестрикции, комплементарны всем хвостам других фрагментов, получаемых с помощью этого же самого рестрикционного фермента.
Таблица 5 - Некоторые рестрикционные ферменты и их сайты рестрикции
При комнатной температуре эти одноцепочечные участки, разных фрагментов, образованных одним рестрикционным ферментом, могут случайно спариться между собой. Их, поэтому, часто называют "липкие концы» ("sticky ends").
Некоторые рестрикционные ферменты, такие как AluI и Smal, разрезают обе цепи ДНК в одной и той же точке в сайте рестрикции, образуя фрагменты с "тупыми" концами ("blunt ends"). На тупых концах таких фрагментов все нуклеотиды обеих цепей ДНК спарены с комплементарными нуклеотидами.
ДНК каждого данного организма имеет совершенно специфический набор сайтов рестрикции. Поэтому, рестрикционные ферменты разрезают данную ДНК на определённый воспроизводимый набор фрагментов, называемых рестрикционными фрагментами.
Рестрикционные ферменты, выделенные из нескольких сотен различных видов бактерий, позволяют разрезать молекулы ДНК на большое число различных фрагментов, соответствующих центрам распознавания этих ферментов (таблица 5).
Названия рестрикционных ферментов обычно пишут курсивом, поскольку они являются сокращениями латинских наименований тех бактерий, в которых они были обнаружены.
9.1.2. Внедрение фрагментов ДНК в векторы. Рестрикционные фрагменты ДНК внедряются в векторы с помощью ДНК-лигаз (рисунок 85). В ходе нормальной репликации ДНК ДНК-лигазы катализируют соединение (склеивание) концов коротких фрагментов ДНК, которые называются фрагментами Оказаки. При клонировании ДНК используются очищенные ДНК-лигазы для ковалентного связывания рестрикционного фрагмента генома и ДНК вектора, имеющих комплементарные концы.
Векторная ДНК и рестрикционный фрагмент ковалентно соединяются вместе посредством стандартных 3'—5' фосфодиэфирных связей ДНК.
Примечательно, что ДНК-лигаза бактериофага Т4, в дополнение к соединению комплементарных липких концов, может соединять любые тупые концы ДНК. Однако соединение тупых концов крайне неэффективно и для такого соединения необходима повышенная концентрация и ДНК, и ДНК-лигазы по сравнению с соединением липких концов.

Рисунок 85 - Сшивание рестрикционных фрагментов с комплементарными липкими концами ферментом Т4 ДНК-лигаза