Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Клонирование ДНК
Векторы на базе бактериофага λ
Векторы, созданные на основе бактериофага λ приблизительно в тысячу раз более эффективнее плазмидных векторов при клонировании большого количества фрагментов ДНК. По этой причине векторы на базе фага-λ широко использовались при создании ДНК библиотек - полных коллекций фрагментов ДНК, представляющих геном или мРНК организма.

Рисунок 88 - Плазмидный вектор pBR322
Два главных фактора объясняют большую эффективность фага λ в качестве клонирующего вектора:
1) заражение клеток Е. coli λ-вирионами происходит в тысячу раз чаще, чем трансформация плазмидами,
2) гораздо большее число λ-клонов, чем трансформированных колоний, может быть выращено и идентифицировано на культуральной среде в чашке Петри.
Когда λ-вирион инфицирует клетку Е. coli, он проходит цикл литического роста, в ходе которого ДНК фаг реплицируется и ассемблируется в более чем 100 дочерних фагов, которые высвобождаются при лизисе инфицированной клетки (см., например, рисунок 69). Если λ-фаги поместить на культуру Е. coli в чашке Петри, то каждый вирион инфицирует единственную клетку. Последующие циклы роста фага приведут к образованию видимых чистых (от клеток) участков, называемых бляшками, в которых разрушены клетки Е. coli и высвобождены дочерние вирионы фагов (см. рисунок 67).
Вирион фага X состоит из головки, заключающей в себе ДНК геном фага, и хвоста, через который происходит инфицирование клеток Е. coli. Гены фага λ, которые кодируют белки головки и хвоста, а также гены, кодирующие различные другие белки, участвующие в репликации ДНК и лизисе клетки, сгруппированы в дискретные участки вирального генома, общая длина которого 49 kb (рисунок 89).
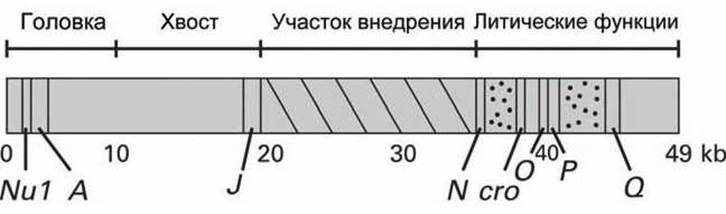
Рисунок 89 - Схема генома бактериофага λ
Однако центральная часть генома фага λ. содержит гены, которые не существенны для литического цикла размножения. Если удалить эти гены и заменить их на другие фрагменты ДНК, то мы получим рекомбинантную ДНК, которая может быть упакована in vitro так, что образуется фаг, способный заражать клетки и формировать бляшки в культуре клеток (рисунок 90).
Для такой in vitro упаковки рекомбинантной ДНК, имитирующей процесс ассемблирования in vivo, необходимы преассемблированные головки и хвосты, и два белка вирусных головок Nul и А.
В ходе поздних стадий инфицирования фагом λ образуется длинная молекула ДНК, называемая конкатомером, которая состоит из множественных копий 49-кb-генома фага, присоединенных друг за другом и разделенных сайтами COS - последовательностями нуклеотидов (которые встречаются в одном геноме только один раз) и к которым присоединяются белки Nul и А для ассемблирования вириона. Присоединение белков Nul и А к сайту COS стимулирует введение внутрь пустой головки сегмента ДНК, лежащего между двумя сайтами COS. После того, как головка заполнена ДНК к ней присоединяется предварительно ассемблированный хвост, в результате чего образуется λ-вириоп, способный инфицировать клетку Е. coli.
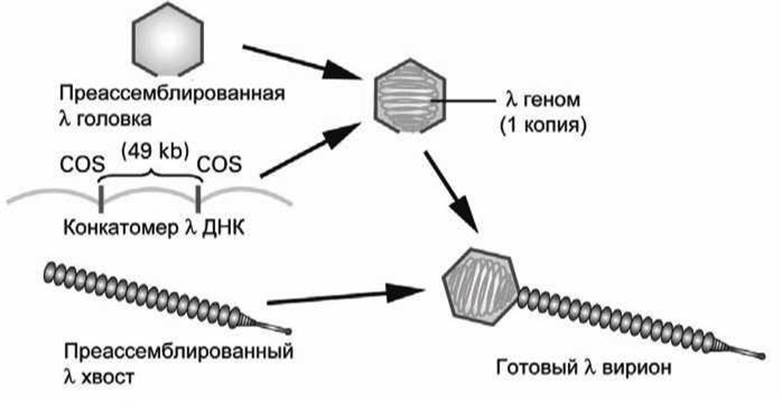
Рисунок 90 - Ассемблирование вириона бактериофага λ
Такие λ-фаги удобно использовать для создания геномной библиотеки - коллекции λ-клопов, которые совместно представляют все ДНК-последовательности генома определённого организма.
Однако при создании таких геномных библиотек для высших эукариот возникают существенные экспериментальные трудности.
Во-первых, гены таких организмов обычно содержат интроны и, поэтому, они слишком длинные, чтобы поместиться целиком внутрь вектора λ-фага. Поэтому последовательности индивидуальных генов разъединены на части и содержатся внутри более чем одного λ-клона (это, кстати, характерно и для плазмидных клонов).
Кроме того, наличие интронов и длинных межгенных участков в геномной ДНК эукариот часто затрудняет идентификацию именно тех существенных частей гена, которые действительно кодируют белковые последовательности.
Поэтому для многих исследований, в качестве исходного материала для создания ДНК библиотеки, более полезной оказывается мРНК, в которой нет некодирующих участков, характерных для геномной ДНК.
В этом случае синтезируются и затем клонируются в векторах фагов ДНК копии с мРНК, которые называются комплементарными ДНК (кДНК, complementary DNA, cDNA). Большая коллекция полученных таким образом кДНК клонов, представляющих все мРНК, экспрессируемые в клетке, называется кДНК библиотекой (cDNA library).