Основы молекулярной биологии. Часть 2: Молекулярные генетические механизмы - А.Н. Огурцов 2011
Транскрипция ДНК
Процессинг РНК эукариот
В прокариотических клетках, не имеющих ядра, трансляция мРНК может начинаться со свободного 5'-конца мРНК в то время, когда 3'-конец все ещё наращивается РНК-полимеразой. То есть транскрипция и трансляция в прокариотах могут происходить одновременно.
В эукариотических клетках не только пространственно разделены транскрипция в ядре и трансляция в цитоплазме клетки, но и первичный транскрипт (про-мРНК) должен быть подвергнут ряду модификаций, который называется процессингом РНК, с тем, чтобы получить функционально зрелую мРНК (рисунок 1, процесс 2). Только после этого мРНК
транспортируется в цитоплазму из ядра. Таким образом, транскрипция и трансляция не могут происходить одновременно в клетках эукариот.
Процессинг мРНК включает в себя несколько операций. В дополнение к уже упомянутому сплайсингу, которому подвергается 95% мРНК, обязательными являются копирование мРНК (100%) (рисунок 15) и полиаденилирование (95%) мРНК.
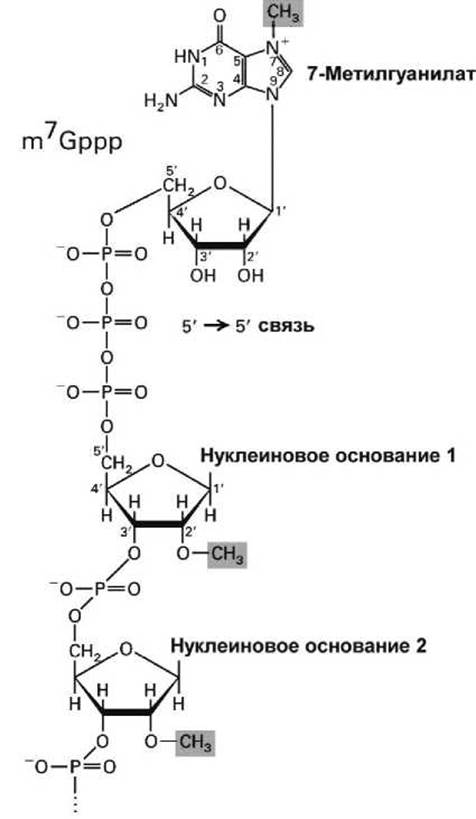
Рисунок 15 - Схема кэпа эукариотической мРНК
Во всех эукариотических про-мРНК немедленно после синтеза оба конца полинуклеотидной цепочки модифицируются.
Процессинг 5'-конца - кэпирование — заключается в том, что как только 5'-конец растущей РНК выходит из РНК-полимеразы, он немедленно кэпируется особыми ферментами.
Кэп (от англ. cap, "шапочка") образуют две операции (рисунок 15):
1) присоединение метилированного гуанозинтрифосфата (ГТФ) (7-Метилгуанилат) в необычной 5'—5' позиции к 5'-концу мРНК;
2) метилирование двух рибоз в первых двух нуклеотидах мРНК. (При этом, однако, у дрожжей не метилируется рибоза первого нуклеотида, а рибоза второго нуклеотида метилируется только у позвоночных).
Функции кэпа:
1) кэп защищает мРНК от ферментативной деградации экзо-нуклеазами;
2) кэп способствует экспорту мРНК в цитоплазму;
3) к кэпу присоединяется фактор трансляции - белок, необходимый для правильной установки мРНК на рибосоме.
Процессинг 3'-конца про-мРНК - полиаденилирование - заключается в разрезании её специфической эндонуклеазой вблизи поли(А)- точки — ААУАА-последовательности и присоединении к освободившемуся 3'-ОН концу поли(А)-цепочки из 100-250 адениннуклеотидов с помощью фермента поли(А)-полимеразы. Длина этой цепочки у дрожжей и беспозвоночных меньше, чем у позвоночных и, в значительной степени, время жизни мРНК коррелирует с длиной поли(А)-хвоста.
Поли(А)-полимераза это составная часть комплекса белков, которые (1) обнаруживают специфическую последовательность в транскрипте, (2) разрезают транскрипт и (3) присоединяют к транскрипту правильное число аденозинфосфатов, осуществляя безматричный синтез.
Финальный этап процессинга про-мРНК - это упоминавшийся выше сплайсинг.
На рисунке 16 представлены основные этапы процессинга эукариотической мРНК, кодирующей трансляцию гена ß-глобина, состоящего из 147 аминокислот. Два интрона прерывают белок-кодирующую последовательность между кодонами для аминокислот 31 и 32, и 105 и 106.
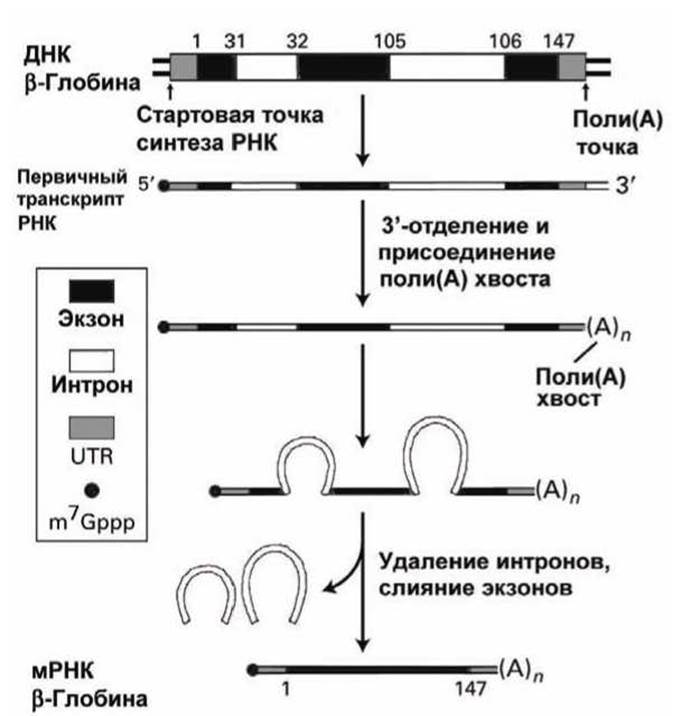
Рисунок 16 - Схема процессинга мРНК на примере гена ß-глобина, который содержит три экзона и два интрона. Кэп m7Gppp обозначен (•)
Транскрипция эукариотического гена начинается до последовательности, которая кодирует первую аминокислоту и продолжается за пределы последнего кодона, в результате чего на концах про-мРНК находятся некодирующие участки, с которых не осуществляется трансляция. Такие нетранслируемые участки (untranslated regions, UTR) не удаляются в ходе процессинга.
5'-Кэп (7-метилгуанозинтрифосфат, m7Gppp) присоединяется в ходе синтеза первичного РНК-транскрипта, который продолжается за пределы поли(А) точки - точки, в которой эндонуклеаза затем разрезает транскрипт, формируя 3'-конец. После присоединения поли(А)-хвоста, (А)n, сплайсинг удаляет интроны и соединяет экзоны.
Удаление интронов происходит в ядре. В про-МРНК два основания, которые непосредственно следуют за экзоном, всегда GU, а последние два основания в интроне, всегда AG. В процессе сплайсинга участвуют несколько малых ядерных РНК, мяРНК (snRNA, small nuclear RNA). мяРНК объединяются с необходимыми белками и формируют сплайсосому. Одна из мяРНК комплементарна к обоим концам интрона. Связывание этой мяРНК с про-мРНК приводит к образованию интронной петли (рисунок 17).
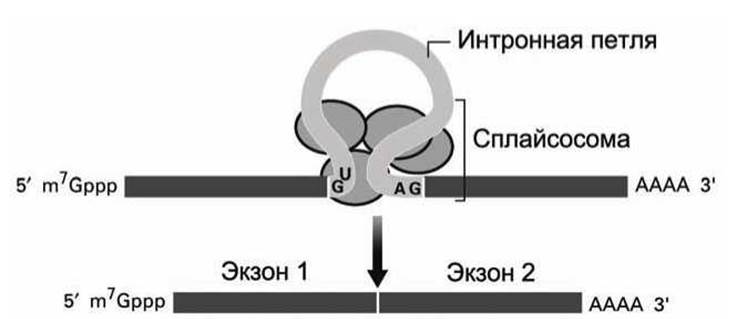
Рисунок 17 - Сплайсинг мРНК
Белки, входящие в состав сплайсосомы, вырезают интрон из про-мРНК и соединяют экзоны. На этом процессинг мРНК заканчивается.
Нетранслируемые участки UTR эукариот достаточно длинные: в клетках млекопитающих длина 5'UTR более сотни нуклеотидов, a 3'UTR - несколько kb (kilobases). мРНК прокариот тоже имеют 5' и 3' UTR-ы, но они обычно содержат менее 10 нуклеотидов.
Наличие интронов в генах многоклеточных эукариот позволяет объяснить синтез многодоменных белков. Если отдельный домен мультидоменного белка кодируется одним экзоном, расположенными между двумя нитронами, то мультидоменная структура белка обеспечивается многократным повторением этого экзона в структуре ДНК, в результате чего цепочка повторяющихся экзонов будет лежать между двумя данными нитронами, обеспечивая мультидоменное повторение.