Основы молекулярной биологии - В.И. Резяпкин 2009
Регуляция экспрессии генов
Регуляция экспрессии генов на уровне организации ДНК
Количество генов в геноме живых организмов составляет у вирусов от нескольких штук до нескольких сотен, у прокариот - от нескольких сот до нескольких тысяч, у эукариот - до нескольких десятков тысяч. Продукты генов нужны различным клеткам в разное время и в неодинаковых количествах. Одни гены в определенный момент развития организма экспрессируются с высокой интенсивностью, другие гены могут находиться в репрессированном состоянии. Очевидно, что экспрессия генов в клетке должна быть согласованной. В связи с этим в клетке существуют тонкие механизмы ее регуляции. Регуляция экспрессии генов может осуществляться на различных уровнях:
✵ на уровне организации ДНК;
✵ на уровне транскрипции;
✵ на уровне созревания РНК;
✵ на уровне деградации РІЖ;
✵ на уровне трансляции;
✵ на уровне посттрансляционных модификаций;
Далее рассмотрим подробнее регуляцию экспрессии генов на каждом уровне.
В зависимости от степени компактизации ДНК ген может находиться в активном или неактивном состоянии. В ядре эукариотической клетки ДНК находится в комплексе с белками, образуя - хроматин. Различают два типа хроматина:
✵ гетерохроматин - компактный хроматин - транскрипционно неактивен;
✵ эухроматин - деконденсированный хроматин - транскрипционно активен.
Активность хроматина пропорциональна его компактности. При активации хроматина происходит его деконденсация.
Регуляция активности генов обусловленная метилированием ДНК
Метилирование ДНК осуществляется в результате обратимого метилирования цитозина, катализируемого ДНК-метилтрансферазой:
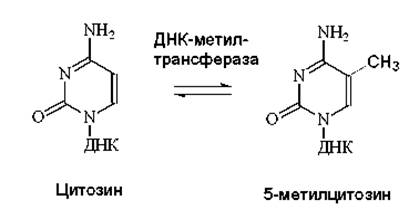
На рисунке 8.1 представлен механизм действия ДНК-метилтрансферазы. После взаимодействия фермента происходит разрыв водородных связей между цитозином и комплементарным ему основанием гуанином. Затем метилтрансфераза присоединяет метильную группу к цитозину. И далее восстанавливаются водородные связи между метилцитозином и гуанином.

Рис. 8.1. Механизм действия ДНК-метилтрансферазы
Не все остатки цитозина в ДНК метилируются. Метилированию подвергаются только те остатки цитозина, которые находятся рядом с гуанинином - ЦГ.
Метилирование ДНК осуществляется после репликации, поэтому сразу же после удвоения молекула ДНК является полуметилированной - одна цепь метилирована, другая - неметилированная. Через определенное время ДНК-метилтрансфераза восстанавливает исходный рисунок метилирования (рис .8.2).
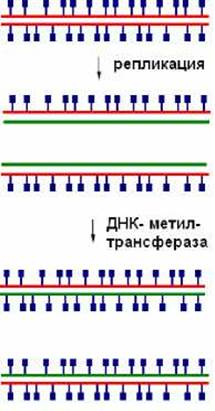
Рис. 8.2. После репликации ДНК-метилтрансфераза восстанавливает исходный рисунок метилирования
Метилирование необходимо для нормального развития организма. Если инактивировать ген метилтрансферазы у мышей, то развитие эмбриона приостановится на ранних стадиях развития. Тем не менее, наличие в составе ДНК метилцитозина опасно для организма, так как при его дезаминировании образуется тимин:
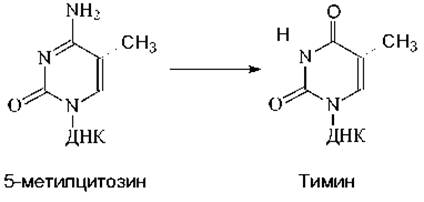
Образовавшийся тимин не комплементарен азотистому основанию гуанину, расположенному на противоположной цепи ДНК. В результате в ДНК образуются неспаренные основания и при ее репликации в одной из двух дочерних молекул ДНК пара ЦГ заменится на ТА, т.е. произойдет мутация (рис. 8.3).
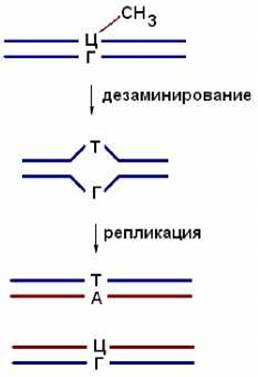
Рис. 8.3. Дезаминирование метилцитозина может привести к мутации
В геноме млекопитающих ЦГ-пары образуют островки, которые часто встречаются в районе промоторов. Метилирование промоторов препятствует образованию транскрипционного комплекса. При этом степень репрессии гена пропорциональна степени метилирования цитозинов. В некоторых случаях метилирование может препятствовать связыванию репрессорного белка с сайленсером. В результате произойдет усиление экспрессии гена.