Основы молекулярной биологии - В.И. Резяпкин 2009
Регуляция экспрессии генов
Регуляция экспрессии генов на уровне транскрипции
Прокариоты
Транскрипция у прокариот может регулироваться на стадиях инициации, элонгации и терминации. Наиболее часто транскрипция регулируется на стадии инициации транскрипции. Регуляция на уровне инициации транскрипции
Эффективность инициации транскрипции зависит в значительной степени от силы промотора. Чем сильнее промотор, тем эффективнее осуществляется синтез РНК. Специфичность связывание РНК-полимеразы прокариот с промотором определяет σ-субъединица. В бактериальных клетках имеется несколько различных σ-субъединиц, обеспечивающих транскрипцию с различных генов. σ-Субъединица необходима только для узнавания промотора, в синтезе РНК она не участвует. Промоторы, узнаваемые одной и той же σ-субъединицей, могут незначительно отличаться друг от друга по первичной структуре. Такие различия и определяют силу промотора. Интересно, что чем ближе структура природного промотора к структуре канонического промотора, тем сильнее промотор.
В клетках прокариот существуют белки регулирующие эффективность транскрипции: белки-репрессоры, блокирующие синтез РНК, и белки-активаторы, положительно влияющие на синтез РНК.
У прокариот процессы транскрипции и трансляции сопряжены, синтез РНК еще не завершился, а рибосомы уже начали синтезировать полипептид (рис. 8.4). Интересно, успешная трансляция способствует транскрипции. Рибосома как бы подталкивает РНК-полимеразу. При отсутствии трансляции синтез РНК может быть заблокирован. Специальный белок «выдергивает» образующуюся РНК.
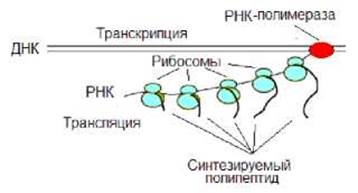
Рис. 8.4. У прокариот процессы транскрипции и трансляции сопряжены
Активность многих промоторов регулируется с помощью особых белков-регуляторов - репрессоров и активаторов. Репрессоры, взаимодействуя с ДНК, снижают активность РНК- полимеразы. В этом случае речь идет о негативной регуляции транскрипции. Участок связывания ДНК с репрессором называется оператором. Существуют низкомолекулярные вещества, влияющие на сродство репрессора к оператору - эффекторы. Различают два вида эффекторов - индукторы и корепрессоры. Индукторы снижают сродство репрессора к оператору, корепрессоры увеличивают сродство репрессора к оператору. Активаторы, напротив, усиливают активность РНК-полимеразы, таким образом, осуществляя позитивную регуляцию экспрессии генов. Некоторые белки могут выступать в одних случаях в качестве репрессора, в других - в качестве активатора.
Далее рассмотрим регуляцию работы лактозного и трип- тофанового оперонов E.coli.
Лактозный оперон (lac-оперон) E.coli
Данный оперон кодирует белки катаболизма лактозы, менее предпочтительного по сравнению с глюкозой источника энергии для данной бактерии. В связи с этим транскрипция lac- оперона происходит только в том случае, если в среде отсутствует глюкоза и присутствует лактоза. В состав lac-оперона входят (рис. 8.5): три структурных гена, кодирующих белки, участвующих в метаболизме лактозы: ß-галактозидазу (гидролизует лактозу), пермеазу (ответственна за транспорт галактозы в клетку), белок А (ß-галактозидтрансацетилаза - функция до конца не выяснена); промотор (40 п.н.); оператор (27 п.н.); область связывания комплекса белка САР (белок активатор катаболитических оперонов) с цАМФ - САР*цАМФ (38 п.н.).
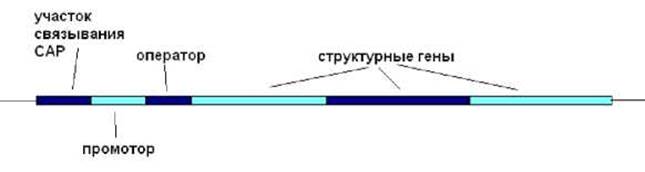
Рис. 8.5. Организация lac-оперона
Транскрипция lac-оперона зависит от наличия или отсутствия лактозы и глюкозы в среде на которой растут клетки E.coli. Рассмотрим три ситуации.
1) В среде, на которой растут клетки E.coli, отсутствует лактоза и присутствует глюкоза.
В результате транскрипции гена репрессора образуется иРНК, трансляция, которой приводит к синтезу белка-репрессора. Последний, взаимодействуя с оператором, не дает
РНК-полимеразе, начать синтез РНК (рис. 8.6). В этом случае белки, ответственные за утилизацию лактозы, не синтезируются. Что вполне логично, если отсутствует лактоза, то и нет смысла в синтезе белков ее утилизирующих.
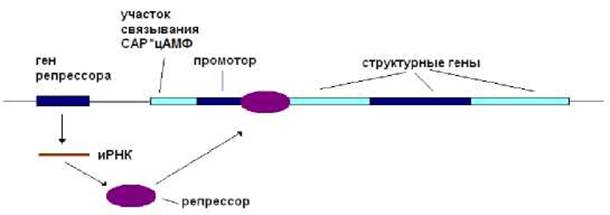
Рис. 8.6. В среде в отсутствие лактозы репрессор, взаимодействуя с оператором, не дает РНК-полимеразе, начать синтез РНК
2) В среде, на которой растут клетки E.coli, присутствует лактоза и отсутствует глюкоза Лактоза, проникнув в бактериальную клетку, связывается с репрессором (рис. 8.7А). Образовавшийся комплекс репрессор-лактоза теряет сродство к оператору и покидает его (рис. 8.7 Б). При недостатке глюкозы в клетке начинает накапливаться цАМФ, которая, связываясь с белком CAP, образует комплекс САР*цАМФ (рис. 8.7 В). Последний взаимодействуя с соответствующим участком ДНК, способствует РНК-полимеразе начать транскрипцию лактозного оперона (рис. 8.7 Г). В результате синтезируется иРНК, которая используется в качестве матрицы для синтеза белков (рис. 8.7 Д), ответственных за катаболизм лактозы. Теперь клетка способна использовать лактозу в качестве источника энергии. Следует отметить, что транскрипция lac- оперона происходит эффективно, когда в среде отсутствует глюкоза и присутствует лактоза.

Рис. 8.7. Транскрипция lac-оперона происходит только в отсутствие глюкозы и в присутствии лактозы
3) В среде, на которой растут клетки E.coli, присутствует лактоза и глюкоза
В этом случае оператор не связан с репрессором, поскольку последний находится в комплексе с лактозой и не обладает сродством к оператору. В тоже время РНК-полимераза не может эффективно осуществлять синтез РНК, так как в клетке концентрация цАМФ недостаточна для образования комплекса САР*цАМФ, необходимого эффективной транскрипции. Таким образом, в присутствии глюкозы и лактозы lac-оперон практически не транскрибируется и не синтезируются белки катаболизма лактозы. Клетка в этом случае предпочитает использовать в качестве источника энергии глюкозу, но не лактозу. Триптофановый оперон (trp-оперон) E.coli
Триптофановый оперон содержит 5 генов, ответственных за синтез триптофана, промотор, оператор (рис. 8.8).
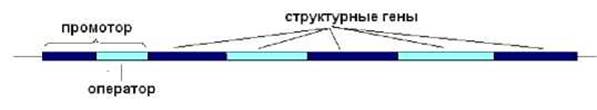
Рис. 8.8. Схема организации триптофанового оперона E.coli
Очевидно, что триптофановый оперон должен транскрибироваться при недостаточном содержании триптофана в клетке. При высоком же содержании этой аминокислоты его экспрессия должна быть слабо выраженной или полностью отсутствовать. В клетках E.coli присутствует репрессор, образующийся в результате экспрессии соответствующего гена. При высоком содержании в клетке триптофана этот репрессор образует комплекс с данной аминокислотой. Образовавшийся комплекс обладает сродством к оператору и, взаимодействуя с ним, не позволяет РНК-полимеразе начать синтез РНК (рис. 8.9). Белки, ответственные за синтез триптофана, не образуются. Соответственно не происходит и синтез аминокислоты.
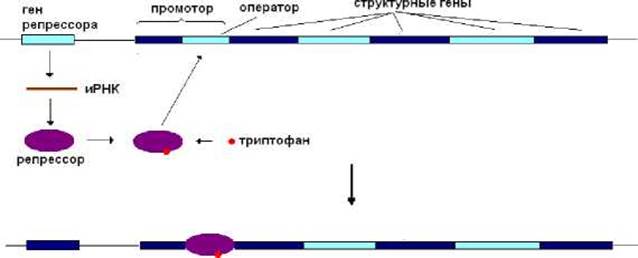
Рис. 8.9. При высоком содержании триптофана в клетке триптофановый оперон не транскрибируется
Как только содержание триптофана в клетке снижается, комплекс триптофан-репрессор распадается. В результате репрессор теряет сродство к оперону. РНК-полимераза приобретает доступ к промотору и начинает транскрипцию структурных генов. Синтезируется иРНК, которая транслируется с образованием белков, ответственных за синтез триптофана. Концентрация этой аминокислоты в клетке постепенно возрастает (рис. 8.10).
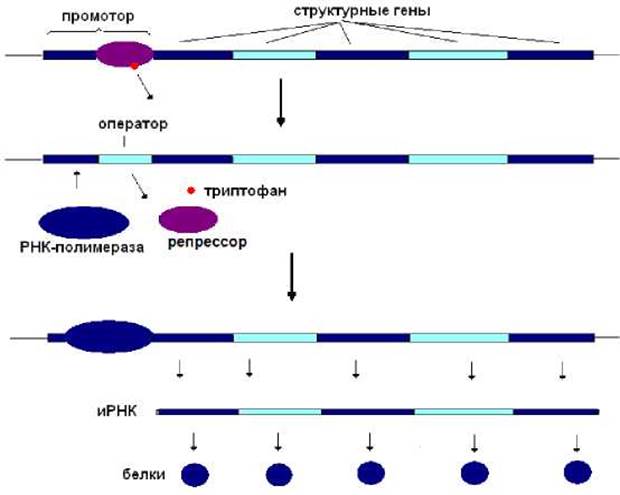
Рис. 8.10. В отсутствие триптофана trp-оперон экспрессируется
Регуляция транскрипции на уровне элонгации Аттенюация
Нами ранее рассмотрена регуляция триптофанового оперона посредством белка-репрессора. В тоже время регуляция этого оперона может осуществляться также благодаря наличию в нем аттенюаторной последовательности (аттенюатора). Эта последовательность входит в состав лидерной области, в которой закодирован также лидерный пептид (рис. 8.11). Лидерный пептид в 10 и 11 положениях содержит два остатка триптофана.

Рис. 8.11. Структура лидерной области триптофанового оперона
Транскрипция trp-оперона может быть преждевременно прервана в области аттенюатора или нет. Преждевременная терминация наблюдается при высоком содержании триптофана в клетке. Так как наличие большого количества аминокислоты не требует ее дополнительного синтеза и, соответственно, ферментов, ее синтезирующих. Напротив, при низких концентрациях триптофана транскрипция оперона осуществляется полностью (рис. 8.12). В итоге происходит синтез иРНК, в результате трансляции которой образуются ферменты, обеспечивающие синтез данной аминокислоты.

Рис. 8.12. При высоких концентрациях триптофана транскрипция trp-оперона преждевременно прерывается в области аттенюатора, а в отсутствие триптофана - осуществляется полностью
Трансляция лидерного участка иРНК сопряжена с транскрипцией. Скорость его трансляции зависит от содержания триптофана в клетке. При высоких концентрациях данной аминокислоты рибосома быстро транслирует этот участок иРНК.
Высокая скорость трансляции способствует формированию структур, приводящих к преждевременной терминации транскрипции. Если рибосома же медленно транслирует лидерный участок иРНК (при низких концентрациях триптофана), то формируется структура, способствующая продолжению транскрипции. В результате синтезируется иРНК, при трансляции которой образуются белки, ответственные за синтез триптофана Регуляция на уровне терминации
Как правило, синтез РНК заканчивается в области терминатора. Однако в некоторых случаях РНК-полимераза не замечает терминатор и прочитывает его, продолжая синтез РНК (рис. 8.13) за его пределами. Этот процесс получил название - антитерминация. Антитерминацию осуществляют специальные белки-антитерминаторы. В результате антитерминации транскрибируются гены, расположенные за терминатором.

Рис. 8.13. Антитерминация. А - синтез РНК завершается в области терминатора. Б - РНК-полимераза при участии белков-антитерминаторов транскрибирует гены, расположенные за терминатором. Р - промотор, Т - терминатор.
Эукариоты
Эффективность экспрессии генов на уровне транскрипции у эукариот зависит от различных факторов. Роль промоторов, энхансеров, сайленсеров, факторов транскрипции рассматривалась ранее в разделе «Транскрипция». В этом разделе мы подробнее остановимся на других способах регуляции экспрессии генов.
Влияние стероидных гормонов на экспрессию генов
Стероидные гормоны регулируют многочисленные биологические процессы, протекающие в организме эукариот. Их действие опосредовано через их влияние на генетический аппарат клеток.
Стероидные гормоны гидрофобны, поэтому они способны легко проникать через клеточные мембраны внутрь клетки в цитоплазму. Здесь гормон взаимодействует с белком- рецептором, являющимся по своей сути фактором транскрипции, с образованием гормон-рецепторного комплекса. Образовавшийся комплекс проникает внутрь ядра клетки, где он взаимодействуя с определенными нуклеотидными последовательностями, активирует транскрипцию зависимых от стероидных гормонов генов. В результате их активации происходит синтез иРНК, которая транспортируется в цитоплазму, где и осуществляется ее трансляция с образованием белков (рис. 8.14). Таким образом, стероидные гормоны обеспечивают синтез белков, которые в конечном итоге и определяют их биологическое действие. Надо отметить, гормон-рецепторный комплекс в ряде случаях тормозит экспрессию некоторых генов.

Рис. 8.14. Механизм действия стероидных гормонов
Влияние гормонов щитовидной железы на экспрессию генов
Гормоны щитовидной железы тироксин и трииодтиронин влияют на протекание многочисленных биологических процессов в организмах человека и животных. Эти гормоны являются производными аминокислоты тирозина (рис. 8.15).
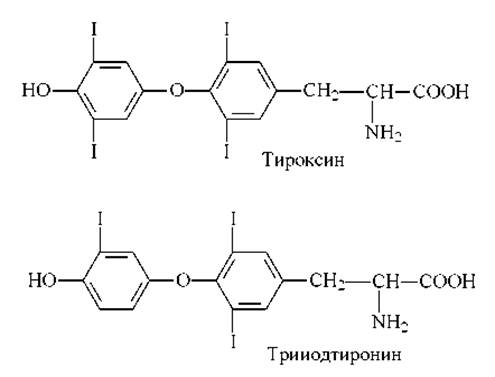
Рис. 8.15. Гормоны щитовидной железы.
На рисунке 8.16 представлен механизм действия тироксина. Этот гормон, так же как и стероидные гормоны, действует на генетическом уровне. Он, проникая через клеточную мембрану, подвергается деиодированию, превращаясь в трииодтиронин. Последний, образуя комплекс с рецептором, активирует транскрипцию определенных генов. Синтезированная иРНК транспортируется из ядра в цитоплазму, где она выполняет роль матрицы для синтеза белка. Вновь образовавшиеся белки обеспечивают гормональный эффект тироксина.
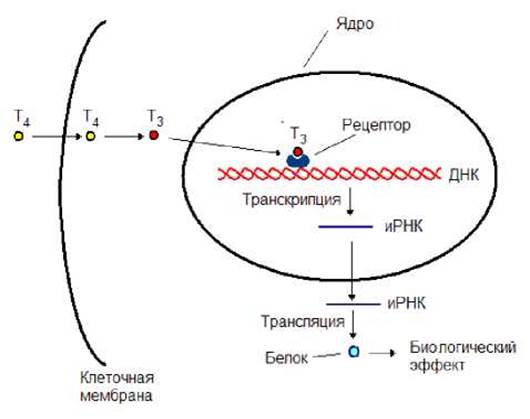
Рис. 8.16. Механизм действия тироксина. Т4 - тироксин, Т3 - трииодтиронин.
Влияние гормонов, рецепторы которых находятся на клеточной мембране, на экспрессию генов
В ядро проникает лишь незначительная часть гормонов: стероидные гормоны и гормоны щитовидной железы. Большинство же гормонов регулируют экспрессию генов, не проникая внутрь клетки. Они оказывают свое действие на генетический аппарат опосредованно через рецепторы, расположенные на плазматической мембране. Известно несколько вариантов передачи сигнала от гормона через рецепторы в ядро.
При первом варианте (рис. 8.17) после взаимодействия гормона с рецептором происходит активация протеинкиназы - фермента осуществляющего фосфорилирование белков (присоединение фосфатной группы к остаткам амнокислот в молекуле белка). В качестве донора фосфатной группы протеинкиназы используют АТФ:
![]()
Активированная протеинкиназа проникает в ядро, где она фосфорилируют один или несколько внутриядерных факторов транскрипции. В результате фосфорилирования изменяется сродство факторов к ДНК и их активность. Что приводит к активации определенных генов. Происходит синтез иРНК. Последняя транспортируется в цитоплазму, где она выполняет роль матрицы для синтеза других факторов транскрипции, которые, проникнув в ядро клетки, запускают более поздние гены. Продукты этих генов и определяют биологический эффект гормонов (рис. 8.17).
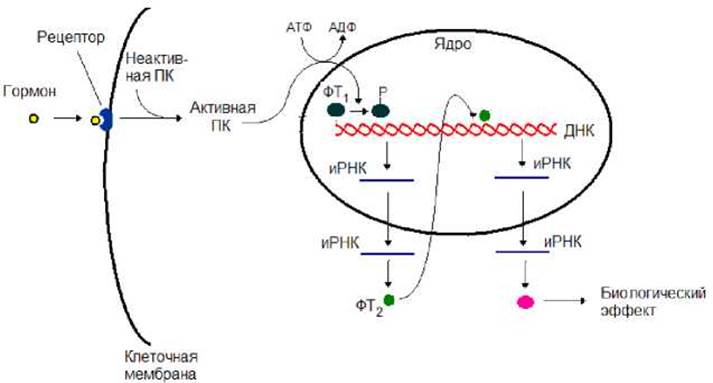
Рис. 8.17. Сигнал от гормона через рецептор в генетический аппарат клетки проникнув в ядро передает протеинкиназа через фосфорилирование внутриядерного фактора транскрипции. ПК - протеинкиназа, ФТ - фактор транскрипции, Р - фосфатная группа.
При втором варианте (рис. 8.18) гормон опосредованно через рецептор активирует протеинкиназу, которая фосфорилирует цитоплазматический фактор транскрипции. Последний после фосфорилирования проникает в клеточное ядро, связывается с ДНК и запускает серию событий, аналогичных рассмотренным выше.

Рис. 8.18. Сигнал от гормона в генетический аппарат клетки через рецептор передает фосфорилированный протеинкиназой фактор транскрипции. ПК - протеинкиназа, ФТ - фактор транскрипции, Р - фосфатная группа.
При третьем варианте после взаимодействия гормона с рецептором протеинкиназа осуществляет фосфорилирование ингибиторной субъединицы в белковом комплексе ингибитор-фактор транскрипции (рис. 8.19). В результате такого фосфорилирования комплекс распадается. Освободившийся фактор транскрипции проникает в ядро и, связавшись с ДНК, запускает серию событий, в результате которых в клетке наблюдается биологический ответ на действие гормона.
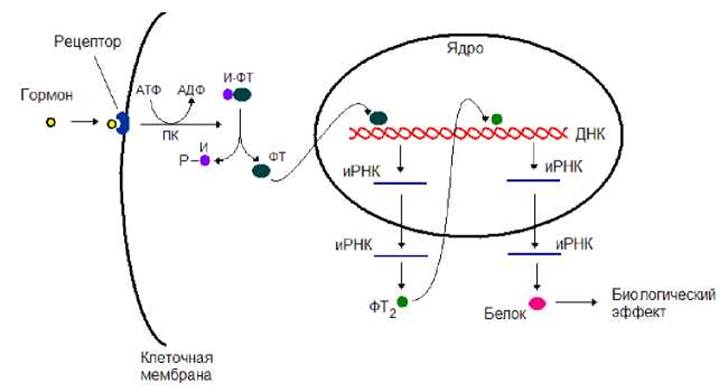
Рис. 8.19. Протеинкиназа, получив сигнал от гормона, фосфорилирует ингибиторную субъединицу в комплексе ингибитор-фактор транскрипции. В результате такого фосфорилирования комплекс распадается. Освободившийся фактор транскрипции проникает в ядро и, связавшись с ДНК, запускает серию событий, в результате которых в клетке наблюдается биологический ответ на действие гормона. ПК - протеинкиназа, ФТ - фактор транскрипции, Р - фосфатная группа, И - ингибитор, И-ФТ - комплекс ингибитор-фактор транскрипции.
Во всех трех рассмотренных вариантах передачи гормонального сигнала в генетический аппарат клетки первичные факторы транскрипци, связываясь с ДНК, запускают транскрипцию ранних генов. Продуктами этих генов являются вторичные факторы транскрипции, которые стимулируют экспрессию поздних генов, определяющих биологический ответ клетки на действие гормона.