Основы молекулярной биологии - В.И. Резяпкин 2009
Регуляция экспрессии генов
Регуляция экспрессии генов на уровне трансляции
Существуют три основных способа регуляции трансляции:
✵ позитивная регуляция трансляции (дискриминация иРНК), основанная на сродстве иРНК к рибосомам и факторам инициации трансляции;
✵ негативная регуляция (трансляционная репрессия), осуществляющаяся с помощью белков-репрессоров, которые, связываясь с иРНК, блокируют инициацию трансляции
✵ тотальная регуляция - регуляция трансляции всей совокупности иРНК.
Позитивная регуляция трансляции (дискриминация иРНК)
У иРНК прокариот для эффективной трансляции инициирующий кодон должен находиться на вершине шпилечной структуры. Предшествовать ему примерно за 3-10 нуклеотидов должна последовательность Шайна-Дальгарно, комплементарная рРНК и обеспечивающая связывание рибосомы в районе инициирующего кодона (рис. 8.25).

Рис. 8.25. У иРНК прокариот инициирующий кодон находится на вершине шпилечной структуры. Ему предшествует за 3-10 нуклеотидов последовательность Шайна-Дальгарно
У эукариот инициация происходит обычно с первого АУГ, однако, если он находится в оптимальном окружении. Эффективность инициации зависит от нуклеотидного состава последовательностей, прилегающих к инициирующему кодону. Наиболее оптимальным для инициации трансляции у млекопитающих является следующая последовательность:
![]()
инициирующий кодон подчеркнут, нуклеотиды, обязательные для инициации выделены жирным. Если первый АУГ находится не в оптимальном контексте, то инициация начинается со следующего АУГ. Для инициации трансляции важным является также наличие на 5’-конце кэпа и на 3’-конце полиА-последовательности. В некоторых иРНК эукариот инициация трансляции осуществляется за счет узнавания внутреннего АУГ. Для такого узнавания требуется протяженная последовательность иРНК, которая узнается специфическими белками, обеспечивающими инициацию с внутреннего кодона.
Частота инициации и скорость трансляции для различных иРНК может значительно отличаться. У прокариот эффективность связывания рибосом зависит от организации рибосомсвязывающих участков иРНК. Чем выше сродство таких участков к рибосоме, тем эффективнее осуществляется инициация трансляции. В связи с этим выделяют «сильные» и «слабые» иРНК (рис. 8.26). На «сильных» иРНК инициация происходит часто, на них рибосомы расположены плотно. В результате синтезируется большое количество молекул белка. На «слабых» иРНК инициация осуществляется реже, рибосомы расположены на значительных расстояниях друг от друга. Синтез белка происходит не столь интенсивно, как в случае «сильных» иРНК.
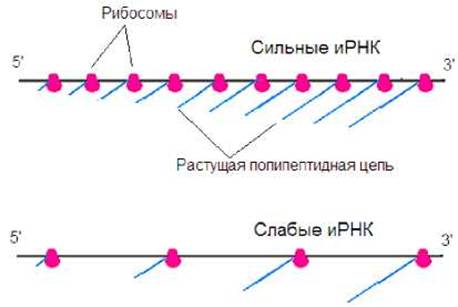
Рис. 8.26. На «сильных» иРНК синтез белка протекает более интенсивно, чем на «слабых» иРНК.
У эукариот иРНК также бывают «слабыми» и «сильными», эффективность инициации трансляции у них обусловлена различным сродством факторов инициации к 5’-концевым структурам иРНК. Сила иРНК определяет количества белка в клетке. Белки, необходимые клетки в больших количествах, кодируются «сильными» иРНК, в малых количествах - «слабыми» иРНК.
У прокариот многие иРНК содержат несколько цистронов. Рибосомы могут инициировать трансляцию цистронов различным образом. В ряде случаях рибосомы осуществляют трансляцию цистронов независимо друг от друга (рис. 8.27А). В других случаях инициация трансляции внутреннего цистрона происходит только после того, как началась трансляции предыдущего цистрона (рис. 8.27Б). В третьих случаях инициация трансляции внутреннего цистрона возможна только после завершения трансляции предыдущего цистрона. При этом большая субъединица рибосомы на границе двух цистронов отделяется, а малая субъединица перемещается на следующий цистрон и обеспечивает инициацию его трансляцию (рис. 8.27В).

Рис. 8.27. Способы трансляции полицистронной иРНК прокариот. А - независимая трансляция цистронов, Б - инициация трансляции внутреннего цистрона происходит только после начала трансляции предыдущего цистрона, В - трансляции внутреннего цистрона возможна только после завершения трансляции предыдущего цистрона, при этом большая субъединица рибосомы на границе двух цистронов отделяется, а малая субъединица перемещается на следующий цистрон.
Негативная регуляция трансляции (трансляционная репрессия)
Негативная регуляция осуществляется при участии репрессорных белков. Последние связываются с иРНК. Часто в месте связывания имеется нестабильная шпилька. Белок- репрессор стабилизирует шпильку, и тем самым препятствует взаимодействию рибосомы с иРНК (Рис. 8.28А) или продвижению рибосомы по иРНК (Рис. 8.28Б). В ряде случаев репрессором является белок кодируемой данной иРНК.
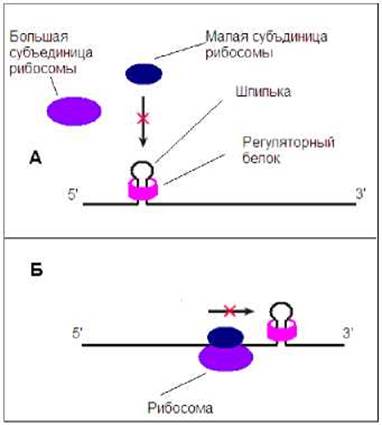
Рис. 8.28. Негативная регуляция трансляции. А - репрессор, стабилизируя шпильку, препятствует взаимодействию рибосомы с иРНК. Б - репрессор, стабилизируя шпильку, препятствует продвижению рибосомы по иРНК.
Рассмотрим в качестве примера регуляцию синтеза ферри- тина (рис. 8.29). Этот белок связывает избыток свободного железа в клетке. Его содержание зависит от концентрации ионов указанного металла. В присутствии железа в клетке происходит синтез ферритина, при его же недостатке белок не синтезируется, поскольку трансляция иРНК ферритина не осуществляется. Трансляция иРНК ферритина зависит от белка-репрессора, который в отсутствии железа связывается с последовательностью, образующую шпилечную структуру на 5’-конце иРНК. Такое связывание стабилизирует шпильку и подавляет трансляцию на стадии инициации. В присутствии ионов железа репрессор образует с ними комплекс и вследствие этого теряет сродство к иРНК. После отделения репрессора иРНК начинает транслироваться, что приводит к накоплению ферритина в клетке. Увеличение содержания ферритина приведет к связыванию железа и снижению концентрации свободного железа в клетке. В результате комплекс репрессора с железом распадается, и репрессор вновь приобретает сродство к иРНК и, взаимодействуя с ней, блокирует ее трансляцию.

Рис. 8.29. Регуляция синтеза ферритина