Основы молекулярной биологии - В.И. Резяпкин 2009
Регуляция экспрессии генов
Регуляция на уровне деградации иРНК
Скорость синтеза каждого конкретного белка зависит от содержания в клетке соответствующей ему иРНК. Чем выше концентрация иРНК, тем, очевидно, и выше скорость синтеза закодированного в ней белка. Количество иРНК в клетке зависит не только от скорости ее синтеза, но и интенсивности ее деградации. Время полужизни иРНК прокариот составляет всего несколько минут, в то время как это показатель у эукариот колеблется от нескольких десятков минут до нескольких суток. В связи с этим роль деградации иРНК в регуляции экспрессии генов у эукариот более выражена, чем прокариот.
В качестве примера рассмотрим значение контролируемой деградации иРНК в регуляции синтеза рецептора трансферина. Регуляция синтеза рецептора трансферта
Скорость поступления железа в клетку зависит от наличия на ее поверхности рецептора трансферина - белка переносчика железа. При недостатке железа в клетке происходит эффективный синтез рецептора трансферина. Вновь синтезированные молекулы рецептора располагаются на клеточной мембране и обеспечивают поступления железа в клетку. При высоком содержании железа в клетке происходит деградация иРНК рецептора трансферина. Что приводит к снижению скорости синтеза рецептора. В результате количество их молекул на мембране уменьшается, что приводит к ослаблению поступления железа в клетку.
Деградация иРНК трансферина определяется шпилечными структурами (элементами нестабильности), расположенными за кодирующими последовательностями (рис. 8.21). Элементы нестабильности связывают регуляторный белок (аконитазу) в форме, не содержащий железо. иРНК рецептора в комплексе с регуляторным белком стабильна и эффективно транслируется. На поверхности клеток увеличивается содержание молекул рецептора трансферина, что приводит к увеличению скорости поступления железа в клетку. При высоком же содержании железа оно связывается с регуляторным белком. Что приводит к ослаблению его сродства к иРНК. Комплекс регуляторный белок- иРНК распадается. иРНК далее подвергается действию РНКаз и деградирует. Снижение количества иРНК приводит к снижению скорости синтеза рецептора. В связи с этим количество его молекул на поверхности клетки уменьшается. Вследствие этого скорость поступления железа в клетку также снижается.
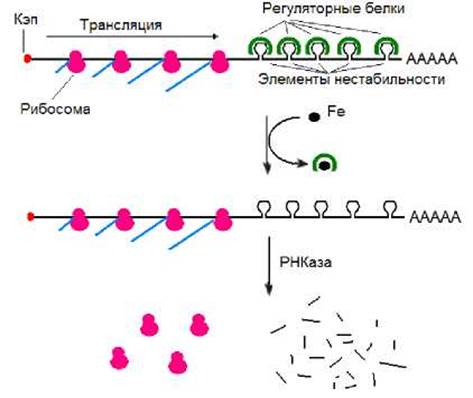
Рис. 8.21. Регуляция синтеза рецептора трансферина
Регуляция экспрессии генов с помощью малых интерферирующих (small interfering) РНК siРНК)
siPHK являются двунитчатыми и состоят из 21 - 28 нуклеотидов. На концах каждой из цепей siPНК имеется два неспаренных нуклеотида. siPНК принимает участие в деградации иРНК и тем самым блокирует синтез белков закодированных в них (рис. 8.22). При появлении siPНК в клетке с ней взаимодействует хеликаза и нуклеаза, образуя комплекс. Хеликаза, используя энергию гидролиза АТФ, раскручивает нити siPНК. Цепь яРНК, к которой присоединена нуклеаза, может связаться с комплементарным участком одноцепочечной иРНК. После этого нуклеаза разрезает ее. Затем частично фрагментированная иРНК подвергается атаке других клеточных РНКаз, которые разрезают ее на более мелкие фрагменты. Таким образом, основной задачей siPНК является деградация тех РНК, которые комплементарны одной из цепочек siPНК.
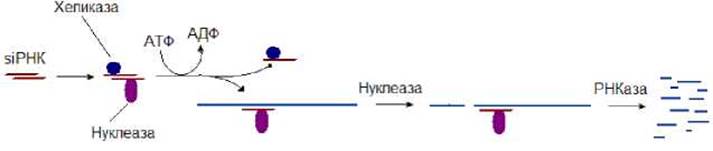
Рис. 8.22. Механизм действия siPHK
siPHK в клетке может образоваться следующим образом. Одна из цепей siPНK присоединившись к цепи иРНК, может с помощью комплекса ферментов, обладающего РНК- зависимой РНК-полимеразной и и РНК-эндонуклеазной активностями, сначала достроить вторую цепь иРНК, а затем разрезать ее с образованием "вторичных" siPНK (рис. 8.23).

Рис. 8.23. Образование siPНK
С помощью siPНK клетка может также подавлять размножение РНК-содержащих вирусов или контролировать перемещение транспозонов по геному.
В медицине siPНK могут использоваться для борьбы с вирусными инфекциями, в генотерапии - для выключения «больных генов».
Маскирование иРНК у экариот
При определенных условиях иРНК может стать недоступной для деградации, инициации трансляции, полиаденилирования 3’-конца. Этот процесс носит название маскирование иРНК. Маскирование иРНК происходит при связывании так называемого маскирующего белка с сегментом маскирования, располагающимся в 3’-некодирующей области иРНК (рис. 8.24). Связывание маскирующего белка с сегментом маскирования приводит к инактивации функций иРНК по всей длине. Маскирование требует наличие и другого менее специфического РНК-связывающего белка, с которым иРНК образует рибонуклеиновый комплекс. Процесс маскирования обратим. Обратный процесс называется демаскированием иРНК.
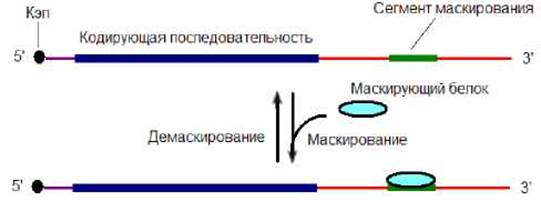
Рис. 8.24. Маскирование-демаскирование иРНК
Маскирование-демаскирование иРНК характерно для многих биологических процессов (гаметогенез, ранние эмбриональное развитие, клеточная дифференцировка и др.). Например в яйцеклетках происходит запасание иРНК в маскированной форме, которая демаскируется после оплодотворения, обеспечивая синтез белка на ранних стадиях развития эмбриона.