Основы молекулярной биологии - В.И. Резяпкин 2009
Перестройки геномов
Сайт-специфическая рекомбинация
Сайт-специфическая рекомбинация не требует протяженных гомологичных участков ДНК. Для ее протекания необходимы определенные короткие гомологичные участки ДНК (1530 п.н.) и специфический ферментативный аппарат. Этот тип рекомбинации характерен для вирусов, прокариот и эукариот. Благодаря сайт-специфической рекомбинации происходит интеграция ДНК умеренных фагов в хромосому бактерий, инверсия определенных участков ДНК в хромосомах бактерий, у высших эукариот при созревании лимфоцитов осуществляются перестройки в последовательностях ДНК, кодирующих иммуноглобулины и др.
Интеграция фага λ в хромосому Е.соli
Первая изученная сайт-специфическая рекомбинация - интеграция фага λ в хромосому E.coli (рис.9.3). Этот процесс достаточно сложен. После проникновения фага λ внутрь клетки E.coli линейная двухцепочечная ДНК переходит в кольцевую форму, благодаря наличию на ее концах комплементарных одноцепочечных последовательностей. Затем ДНК фага интегрирует в хромосому бактерии в строго определенном месте. Фаг, интегрированный в хромосому бактерии, называется профагом. Возможно также и вырезание профага из хромосомы. Этот процесс протекает в обратной последовательности. Для интеграции фага необходимы белок, закодированный в геноме фага, - интеграза - и белок IHF (integratin host factor) бактериального происхождения. Для вырезания профага из хромосомы кроме указанных белков еще дополнительный продукт одного из фаговых генов.

Рис. 9.3. Интеграция фага λ в хромосому Е. coli
Сайт-специфическая рекомбинация у фага Mu
В центре ДНК фага Mu располагается сегмент G, размер которого составляет около 3000 п.н. На его флангах имеются инвертированные повторы (около 30 п.н.). В каждом повторе содержится сайт, участвующий в рекомбинации. Ее осуществляет закодированный в геноме фага фермент - инвертаза, в результате действия которой происходит инверсия G-сегмента (рис. 9.4). Этот процесс является обратимым. При одной ориентации сегмента G транскрибируются гены, ответственные за адсорбцию и размножение фага на одних бактериальных клетках. При другой ориентации сегмента с того же самого промотора транскрибируются другие гены, ответственные за адсорбцию и размножение фага на других бактериальных клетках. Таким образом, инверсии сегмента позволяют фагу Mu менять круг хозяев, в которых он может размножаться.
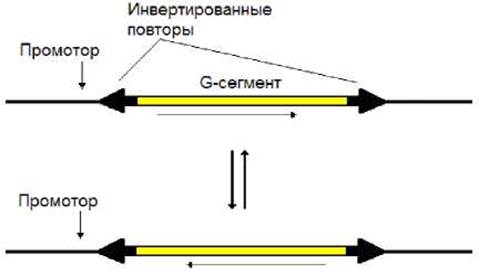
Рис. 9.4. Сайт-специфическая инверсия сегмента Gу фага Mu
Сайт-специфическая рекомбинация у Salmonella typhimurium
Salmonella typhimurium является патогенной бактерией для грызунов. Жгутики этой бактерии построены из белков - флагеллинов. После попадания бактерии в организм животных у последних вырабатываются антитела к флагеллину. Эти антитела совместно с другими компонентами иммунной системы способны обезвредить бактерии. Однако после того как в организме хозяина образовались антитела к данному антигену, у некоторых бактерий происходит переключение генов флагеллинов. Благодаря этому прекращается синтез одного флагеллина и начинается синтез другого флагеллина, и жгутики приобретают другие антигенные свойства. Выработанные ранее антитела станут не эффективными. Такие клетки начнут быстро размножаться в организме хозяина и вызовут новую волну инфекции.
За смену флагеллинов у Salmonella typhimurium ответственен расположенный в хромосоме сегмент Н (993 п.н.). Этот сегмент периодически подвергается обратимым сайтспецифическим инверсиям (рис. 9.5.). Сегмент Н на флангах имеет обращенные повторы. В его пределах располагается ген, кодирующий инвертазу, осуществляющую инверсию сегмента, и два промотора Р1 и Р3. Промотор Р1 обеспечивает синтез транскрипта с генов флагеллина Н2 и репрессора гена флагеллина Н1. В результате трансляции образовавшейся иРНК синтезируются соответственно флагеллин Н2 и репрессор гена флагеллина Н1. Флагеллин Н2 формирует жгутики, а репрессор связывается с оператором гена флагеллина Н1 и блокирует его экспрессию. Промотор Р3 обеспечивает транскрипцию гена инвертазы. Образовавшаяся иРНК служит матрицей для синтеза инвертазы. Последняя катализирует инверсию сегмента Н, в результате промотор Р1 перемещается на значительное расстояние от генов, экспрессию которых он обеспечивал. Вследствие этого гены флагеллина Н2 и репрессора гена флагеллина Н1 перестают экспрессироваться. Соответственно флагеллин Н2 и репрессор гена флагеллина Н1 не образуются. Ген же инвертазы продолжает экспрессироваться. Отсутствие синтеза репрессора приводит к тому, что оператор гена флагеллина Н1 освобождается и с промотора Р2 начинается синтез иРНК, трансляция которой приводит к образованию флагеллина Н1. Так происходит переключение синтеза флагеллинов. Рассмотренная нами сайтспецифическая инверсия у Salmonella typhimurium позволяет бактерии обманывать иммунную систему хозяина.
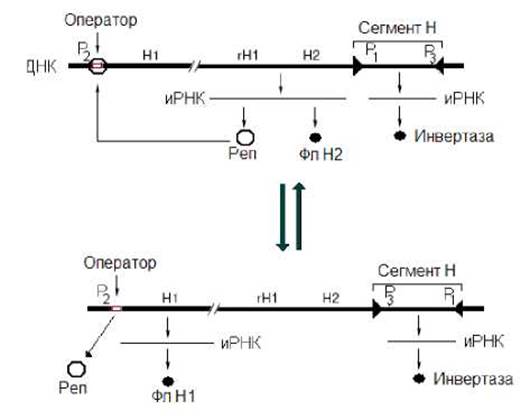
Рис. 9.5. Сайт-специфическая инверсия у Salmonella typhimurium. Н1 - ген флагеллина Н1, Н2 - ген флагеллина Н2, rH1 - ген репрессора гена флагеллина Н1, Фл Н1 - флагеллин Н1, Фл Н2 - флагеллин Н2, Реп - репрессор
Перестройки ДНК у бактерий, обуславливающие воссоединение кодирующих последовательностей
Кодирующие последовательности некоторых генов бактерий прерваны вставками - специфическими ДНК-элементами.
Они на флангах имеют короткие прямые повторы и кодируют сайт-специфическую рекомбиназу. Последняя способна узнавать прямые повторы, вырезать элемент в виде кольцевой молекулы и воссоединять разделенные участки генов (рис. 9.6). Вырезание вставки происходит с абсолютной точностью. Размер вставки может колебаться от 10 т.п.н. до нескольких 10 т.п.н. Благодаря такой перестройки ДНК происходит формирование функционально активного гена.
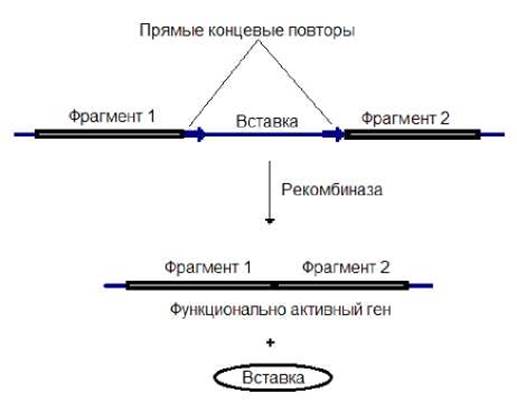
Рис. 9.6. Воссоединение кодирующих последовательностей гена при участи рекомбиназы