Основы молекулярной биологии - В.И. Резяпкин 2009
Мобильные генетические элементы
Ретротранспозоны с длинными концевыми повторами (ДКП)
Ретранспозоны с ДКП состоят из центральной части, называемой «тело», и имеющей размер 5000 - 8000 п.н. На их флангах располагаются прямые ДКН, состоящие обычно из 300 - 400 п.н. В составе ДКП содержатся участки, ответственные за инициацию транскрипции и полиаденилирование. Ограничивают ретротранспозон короткие прямые повторы, возникшие в результате дупликации сайта-мишени при его встраивании (рис. 10.8). Число копий ретротранспозонов, принадлежащих к одному семейству, в геноме варьирует от нескольких до сотен тысяч.

Рис. 10.8. Ретротранспозон с ДКП
Один из способов перемещения ретротранспозона сходен со схемой жизненного цикла ретровирусов. В перемещении по этому механизму принимают участие белки, закодированные в теле МГЭ, - интеграза и обратная транскриптаза (ревертаза). Механизм ретротранспозиции представлен на рис. 10.9. В начале в результате транскрипции ретротранспазона, осуществляемой РНК-полимеразой II, образуется его РНК-копия, которая далее транспортируется из ядра в цитоплазму. Затем РНК при участии обратной транскриптазы используется в качестве матрици для синтеза ДНК-копии мобильного элемента. Последняя транспортируется в ядро клетки и с помощью интегразы встраивается в новый участок ДНК. Так ретротранспозон может распространяться по геному.
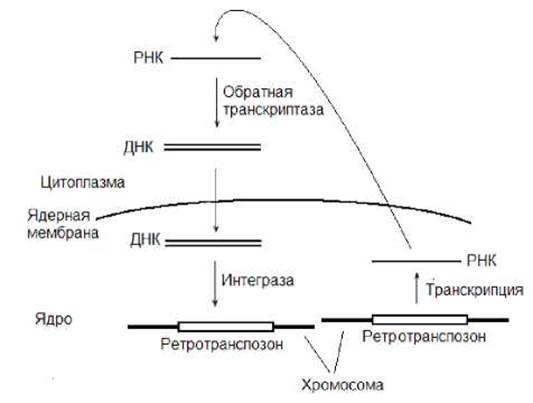
Рис. 10.9. Схема перемещения ретротранспозона, включающая образование РНК-копии

Рис. 10.10. Схема перемещения ретротранспозона с использованием механизма гомологичной рекомбинации
Ретротранспозоны могут также перемещаться, используя механизм гомологичной рекомбинации. При таком перемещении (рис. 10.10) в результате гомологичной рекомбинации между ДКП ретронспозон вырезается из хромосомы, при этом в составе генома остается один ДКН, другой оказывается в составе вырезавшегося МГЭ. ДКП, сохранившийся в геноме, посредством гомологичной рекомбинации, способен обеспечить встраивание в этом участке другого представителя этого же семейства ретротранспозонов, имеющего такую же структуру ДКП.
Ретротранспозоны ответственны за ряд генетических событий. При его внедрении внутрь экзона может произойти инактивация гена (рис. 10.11). Причиной этого является нарушение первичной структуры последнего.
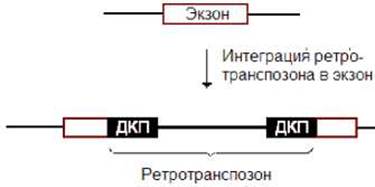
Рис.10.11. Внедрение ретротранспозона в экзон может привести к инактивации гена
Ретротранспозон может интегрировать и в интрон. В связи с этим возможна такая ситуация. ДКП, оказавшись за одним из экзоном, может обеспечить преждевременную терминацию транскрипции благодаря наличию терминатора в его составе. В результате будет синтезированна укороченная молекула РНК. Другой ДКП может обеспечить транскрипцию следующего эк- зона со своего промотора (рис. 10.12). Таким образом, в этом случае внедрение ретротранспозона обуславливает образование двух молекул РНК вместо одной.

Рис. 10.12. Внедрение ретротранспозона в интрон может привести к синтезу двух молекул РНК вместо одной
Внедрение ретротранспозона рядом с геном может усилить его экспрессию за счет энхансера в составе ДКП (рис. 10.13) или за счет сильного промотора в составе ДКП (рис. 10.14).

Рис. 10.13. Внедрение ретротранспозона рядом с геном может усилить его экспрессию за счет энхансера в составе ДКП

Рис. 10.14. Внедрение ретротранспозона рядом с геном может усилить его экспрессию за счет сильного промотора в составе ДКП
Еще одной интересной особенностью ретротранспозонов является их способность залечивать двухцепочечные разрывы ДНК.