Основы молекулярной биологии - В.И. Резяпкин 2009
Организация гемонов
Транскрипция и репликация одноцепочечных (-)РНК геномов
Геном таких вирусов может быть представлен одной или несколькими молекулами (-)РНК. В состав вирусных частиц входит РНК-полимераза, способная использовать (-)РНК в качестве матрицы для синтеза (+)РНК.
Рассмотрим транскрипцию и репликацию вируса везикулярного стоматита (ВВС). Его геном представлен одной молекулой (-)РНК, длина которой составляет около 11 тысяч нуклеотидов. После проникновения вируса в клетку в цитоплазме начинается синтез моноцистронных иРНК, который осуществляется РНК-полимеразой. Этот фермент прочитывает геномную (-)РНК, начиная с ее 3’-конца и перемещаясь в направлении к 5’-концу. Между участками, кодирующими индивидуальные белки, в геномной (-)РНК располагаются полиУ-последовательности, состоящие из 7 нуклеотидов. РНК-полимераза в этой области как бы пробуксовывает, при этом она многократно прочитывает эту последовательность. В результате на 3’-конце иРНК синтезируется полиА-последовательность, образованная несколькими десятками или сотнями нуклеотидов. По окончанию ее синтеза осуществляется терминация транскрипции. Затем происходит реинициация транскрипции на следующем участке (-)РНК. Описанные события осуществляются на границе между всеми генами. В результате образуются иРНК, 5’-концы которых кэпируются (рис. 11.18). После их трансляции и накопления белка начинается репликация вирусного генома. В связи с этим подавляется «буксование» РНК-полимеразы и синтезируются полномерные (+)РНК (антигеном), которые далее копируются РНК-полимеразой с образованием геномной (-)РНК (рис. 11.18).
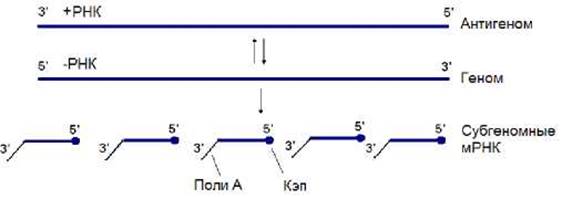
Рис. 11.18. Схема репликации и транскрипции генома ВВС
Геном вируса гриппа является сегментированным и представлен 8 молекулами одноцепочечных (-)РНК от 900 до 2350 нуклеотидов. После проникновения вируса в клетку его геном транскрибируется и происходит синтез вирусной иРНК. При этом каждая молекула (-)РНК копируется не полностью. Это связано с тем, что за 20 нуклеотидов до ее 5’-конца имеется по- лиУ-последовательность, состоящая из 5 - 7 нуклеотидов. На этом участке вирус-специфическая РНК-полимераза «пробуксовывает», многократно прочитывая эту последовательность. В результате на 3’-конце иРНК синтезируется полиА-последовательность (рис. 11.19). По окончании ее синтеза осуществляется терминация транскрипции. Таким образом, синтезированная субгеномная РНК (иРНК) имеет полиА-последовательность и короче геномной (в связи с этим, она не может служить матрицей для синтеза генома). В результате транскрипции синтезируются 8 субгеномных иРНК, часть молекул двух из этих РНК подвергаются сплайсингу. В итоге образуются 10 различных иРНК, используемые для синтеза вирусспецифических белков. После их накопления синтезируются полномерные (+)РНК. Последние служат матрицами для синтеза геномных (-)РНК (рис. 11.19).
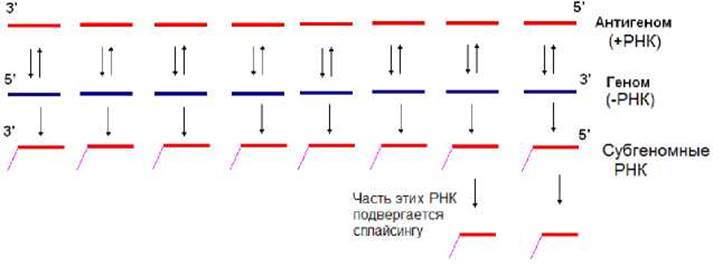
Рис. 11.19. Схема репликации и транскрипции генома вируса гриппа Репликация и транскрипция двуцепочечных РНК-геномов
Двухцепочечные РНК- геномы встречаются как у вирусов прокариот, так и эукариот. Представителем этой группы вирусов являются реовирусы. В их сердцевине содержится 10 сегментов двухцепочечных РНК, РНК-полимераза, способная использовать РНК-дуплекс в качестве матрицы для синтеза (+)РНК. После проникновения в клетку вируса сегменты двухцепочечной РНК используются в качестве матрицы для синтеза (+)РНК, при этом родительская цепь из дуплекса не вытесняется. После синтеза 5’-конец дочерней цепи (+)РНК кэпируется, 3’-конец полиаденилированию не подвергается. Вновь синтезированная (+)РНК на первой стадии служит матрицей для синтеза белка, а затем она копируется РНК-полимеразой, в результате образуется двухцепочечная молекула РНК (рис. 11.20.).

Рис. 11.20. Схема репликации реовирусов