ЗАГАЛЬНА МІКРОБІОЛОГІЯ - Т.П. Пирог - 2004
12. МЕТАБОЛІЧНА АКТИВНІСТЬ АЕРОБНИХ ГЕТЕРОТРОФІВ
12.1. МЕТАБОЛІЗМ С2-СПОЛУК
12.1.1. Етанол та ацетат як субстрати
Окиснення етанолу. Першим етапом метаболізму етанолу у бактерій є його окиснення до ацетальдегіду, яке здійснюється ферментом алкогольдегідрогеназою (рис. 12.1). Відомі два типи алкогольдегідрогеназ: НАД(Ф) - та ПХХ-залежні ферменти. У першому випадку акцептором електронів є НАД або НАДФ , у другому — піролохінолінхінон. ПХХ-залежна алкогольдегідрогеназа виявлена у бактерій роду Pseudomonas. Окиснення етанолу у Acinetobacter calcoaceticus та Acetobacter aceti здійснюється НАД -залежним ферментом. У 90-і роки XX ст. у деяких грампозитивних бактерій виявлено новий тип нікотинпротеїнових алкогольдегідрогеназ, які використовують Ν,Ν -диметил-4-нітрозоанілін як акцептор електронів.
Окиснення ацетальдегіду. Окиснення ацетальдегіду відбувається за участю ферментів ацетальдегіддегідрогеназ. У першому випадку НАД(Ф) -залежні ферменти каталізують окиснення ацетальдегіду до ацетату. У другому випадку НАД -залежний фермент (ацетаильдегіддегідрогеназа ацилювальна) акцептує кофермент А, і з ацетальдегіду утворюється ацетил-КоА (без стадії утворення ацетату). Так, за участю цього ферменту утворюється ацетил-КоА з ацетальдегіду і коферменту А у бактерій Pseudomonas sp.
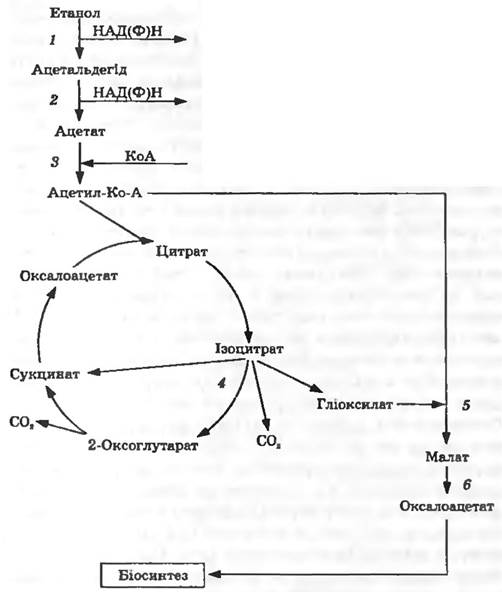
Рис. 12.1. Схема метаболізму етанолу
Метаболізм ацетату. Залучення ацетату до метаболізму відбувається двома шляхами: за участю ацетаткінази та фосфотрансацетилази або за допомогою ацетил КоА-синтетази, яка акцептує КоА з утворенням ацетил-КоА (див. рис. 12.1). Для багатьох бактерій, у тому числі Escherichia coli, характерна наявність як ацетаткінази, так і ацетил-КоА-синтетази, причому E. coli реалізує один з двох шляхів асиміляції ацетату залежно від концентрації цього субстрату в середовищі.
На наступному етапі ацетил-КоА вводиться у цикл трикарбонових кислот (див. рис. 12.1).
Гліоксилатний цикл. Виникає запитання: як під час росту на етанолі чи ацетаті відбувається регенерація проміжних продуктів циклу трикарбонових кислот, необхідних для процесів конструктивного метаболізму? Під час росту на глюкозі такими анашіеротичними реакціями є карбоксилювання пірувату та фосфоенолпірувату. Але під час вирощування на (С2-субстратах ці реакції функціонувати не можуть, оскільки з етанолу та ацетату прямого утворення фосфоенолпірувату не відбувається.
У цьому разі анаплеротичною послідовністю реакцій є гліоксилатний цикл. Його називають також циклом Кребса—Корнберга. Ця анаплеротична послідовність реакцій здійснюється за участю двох ключових ферментів: ізоцитратліази та малатсинтази (див. рис. 12.1). Ізоцитратліаза розщеплює ізоцитрат на сукцинат і гліоксилат, а малатсинтаза каталізує приєднання гліоксилату до ацетил-КоА з утворенням малату. Отже, у гліоксилатному циклі під дією ізоцитратліази та малатсинтази відбувається перетворення одного моля ізоцитрату і одного моля ацетил КоА у два моля С4-дикарбонових кислот (сукцинат і малат), які далі перетворюються наоксилоацетат. Під час росту на етанолі чи ацетаті оксалоацетат є вихідною сполукою для синтезу вуглеводів, необхідних для процесів конструктивного метаболізму.
Глюконеогенез. Дійсно, під час росту на етанолі чи ацетаті клітинам доводиться не тільки поповнювати витрати проміжних продуктів циклу трикарбонових кислот, що відбувається у гліоксилатному циклі, а й синтезувати глюкозу та її похідні, які є необхідними для конструктивного метаболізму (біосинтезу полісахаридів, нуклеїнових кислот та ін.). Ці перетворення здійснюються шляхом глюконеогенезу (рис. 12.2).

Рис. 12.2.- Реакції глюконеогенезу
Оксалоацетат перетворюється нафосфоенолпіруват під дією ферменту фосфоенолпіруваткарбоксикінази, який є ключовим ферментом глюконеогенезу. А далі проходять реакції гліколізу, тільки в зворотному напрямку: з ФЕП утворюється фосфогліцерат (ферменти фосфогліцератфосфомутаза та гліцератфосфомута за), потім 1,3-дифосфогліцерат (фермент фосфогліцераткіназа), триозофосфати — гліцеральдегід-3-фосфат та діоксиацетон (фермент гліцеральдегідфосфатдегідрогеназа) і фруктозо-1,6-дифосфат (фермент фруктозодифосфатальдолаза).
Далі спостерігаються відмінності від гліколізу, зумовлені тим, що у гліколізі реакції, які каталізуються фосфофруктокіназою та гексокіназою, є незворотними. Для їх обходу використовуються інші ферментативні реакції. Так, фруктозо-6-фосфат утворюється з фруктозо-1,6-дифосфату під дією ферменту гексозодифосфатази, а глюкозо-6-фосфат перетворюється на глюкозу в реакції, яка каталізується глюкозо 6-фосфатазою.