Общая микробиология - Шлегель Г. 1987
Микроорганизмы и окружающая среда
Экология микроорганизмов
В предыдущих главах были описаны различные микроорганизмы, сгруппированные по их физиологическим и биохимическим свойствам. Упоминалось и о местах обитания. Полученные сведения позволяют теперь рассмотреть отношения микроорганизмов с окружающей их средой. Мы остановимся сначала на основных понятиях и представлениях экологии. Эта наука изучает поведение организмов в их естественных местообитаниях, взаимоотношения их между собой и с окружающей средой.
Первые следы жизни относятся к периоду более 3 млрд. лет назад; это были микроорганизмы, которые преобладали в биосфере Земли до периода около 0,5 млрд, лет назад. Таким образом, прокариоты не только стоят у истоков земной жизни, из них не только развилось все многообразие эукариотических форм, но они и после этого существовали всегда. Высшие формы жизни на протяжении своей эволюции никогда не были одиноки; их постоянно либо теснили, либо поддерживали вездесущие одноклеточные организмы. Среди современных высших форм жизни есть и такие, которые утвердились не только в борьбе с себе подобными, но и во взаимоотношениях с микроорганизмами. У многих организмов в процессе эволюции выработались терпимые, партнерские отношения - мутуалистический симбиоз. Часть этой главы (разд. 17.2) будет специально посвящена микроорганизмам как симбионтам других микроорганизмов, растений и животных.
Микроорганизмы уже существовали, когда поверхность нашей планеты принимала свой нынешний вид; они уже присутствовали в то время, когда сдвигались континенты, создавались отложения толщиной в несколько тысяч метров, земная кора много раз опускалась и сминалась в складки, возникали залежи руд, угля, месторождения нефти и природного газа. Во многих из этих процессов активно участвовали микроорганизмы (о некоторых вопросах геомикробиологии речь пойдет в разд. 17.3).
На протяжении не менее чем 80% всего периода органической эволюции Земля была населена исключительно микроорганизмами. Если ископаемые остатки микробов находят редко, то данные сравнительной физиологии и биохимии служат достаточной опорой для классификации прокариот по типу метаболизма. Однако при чтении раздела об эволюции организмов следует учитывать то, что в этой области еще много пробелов и домыслов.
17.1.1 Введение
Было приложено немало усилий, чтобы выделить и описать отдельные микроорганизмы. Исследования на чистых культурах имеют существенное значение, так как помогают оценить потенциальные возможности и функции отдельных видов в природе. Таксономические, физиологические, биохимические и генетические исследования некоторое время уводили микробиологов в сторону от основной цели их науки - от изучения поведения микроорганизмов в их естественных местах обитания.
Взаимоотношениями организмов между собой и с окружающей средой занимается экология. Экология микроорганизмов исследует лишь отдельные части целостных экологических систем. Экология - очень разносторонняя наука, и ее проблемы разрабатываются представителями ряда частных биологических дисциплин и рассматриваются в разных аспектах. В связи с этим даже в употреблении основных экологических понятий нет единства. Мы попытаемся сначала пояснить некоторые общие термины, применяемые в экологии микроорганизмов.
Экосистема. Основной единицей в экологии является экосистема. В нее входят как биотические, так и абиотические компоненты. Биотические компоненты составляют сообщество организмов, или биоценоз. При этом речь идет, как правило, о популяциях микроорганизмов, причем популяция может состоять из клонов одного вида или нескольких различных видов. Под абиотическими компонентами следует понимать физические и химические условия экосистемы, в которой живут организмы. Размеры микробных экосистем очень разнообразны. Это может быть, например, пруд, озеро или корневая система дерева. Возможны и такие малые экосистемы, как ротовая полость человека, рубец жвачного животного или участок кишечника. Все жизненное пространство нашей планеты в совокупности - биосферу - можно рассматривать как гигантскую экосистему. С той или иной экосистемой часто связывают понятие «окружающая среда». Эта среда поддерживает взаимоотношения определенного организма (или популяции) с окружающими его (ее) биотическими или абиотическими компонентами экосистемы.
Местообитание. В пределах экосистемы для каждого вида можно описать его местообитание. Это тот участок или жизненное пространство (местность), в котором обычно живет данный организм (индивидуум или популяция). Для каждого организма можно указать как минимум одно местообитание, в котором он обычно встречается, где он благополучно растет и развивается и где его с большой вероятностью можно встретить вновь и выделить. Местообитанием могут быть донные отложения моря, плодородная гумусная почва, носовая полость или кишечник человека. В рамках определенной экосистемы микроорганизм имеет, как правило, только одно-единственное местообитание; но вообще он может иметь и несколько таких мест, каждое в отдельной экосистеме. Например, бактерии рода Rhizobium растут как в почве, так и в растениях; определенная метанобразующая бактерия может обитать в морских отложениях, в рубце жвачных или в метантенке очистной установки. Иными словами, местообитание - это «улица» и «номер дома» данного организма; некоторые организмы могут иметь по нескольку адресов.
Экологическая ниша. В отличие от термина «местообитание» понятие «экологическая ниша» отражает не место в пространстве, а функцию какого-то вида или популяции в сообществе организмов. Экологическая ниша характеризует «профессию» вида. Можно исходить из того, что каждый вид (или популяция) выполняет определенную функцию, которая обусловлена его (ее) потребностями в пище, подвижностью, способом размножения, биохимическими возможностями, структурными особенностями и пределами толерантности к условиям среды. Может или не может какой-либо вид выполнять определенную функцию в определенной экосистеме, зависит от совокупности его свойств. Обычно границы распространения какого-то вида или популяции фактически бывают уже, чем можно было бы ожидать, исходя из свойств этого вида. Иными словами, реальные ниши, как правило, уже потенциальных. Будет ли вид действительно выполнять ту функцию, к которой он потенциально способен,-это часто определяется вторичными, привходящими обстоятельствами.
Поясним это на примере. В рубце жвачных могут расти и выполнять функцию расщепления целлюлозы только те целлюлозолитические бактерии, которые осуществляют это в анаэробных условиях и способны получать энергию в результате брожения. Далее, они должны быть толерантны к температуре внутри желудка, к присутствию жирных кислот, ферментов, аммиака, газов и других продуктов. И наконец, должно быть обеспечено непрерывное удаление определенных продуктов брожения, например Н2. Таким образом, для выполнения данной функции в определенной экосистеме вид должен обладать целым рядом специфических особенностей.
Обитатели экосистемы. Согласно концепции, выдвинутой Виноградским в 1925 г., микроорганизмы, встречающиеся в экосистеме, можно подразделить на две категории - автохтонные и аллохтонные. Автохтонные микроорганизмы являются типичными обитателями данной экосистемы (например, почвы, кишечника) и присутствуют там всегда. Их неизменно можно найти, скажем, в почве независимо от того, вносятся туда извне определенные питательные вещества или нет. Наличие таких видов обусловлено более или менее постоянным присутствием питательных веществ, характерных для данной экосистемы. Под аллохтонными (или зимогенными) микроорганизмами понимают такие, наличие которых зависит от случайного повышения концентрации питательных веществ или от добавления определенных веществ. Такие виды в известной мере чужды данной экосистеме, присутствуют в ней временно или пребывают в состоянии покоя.
К автохтонным обитателям экосистем относятся обычно высокоспедиализированные организмы, такие как нитрифицирующие бактерии, обитатели горячих источников и других экстремальных экосистем. К зимогенным представителям принадлежат многие вездесущие почвенные и водные бактерии (убиквисты).
Численность и разнообразие микроорганизмов в экосистемах. В нормальных условиях в почве или в воде развивается большое число видов. «Нормальными» условиями мы считаем нейтральное значение pH, обилие питательных веществ и высокое содержание воды. Чем больше условия в экосистеме отклоняются от нормальных, т. е. чем более экстремальными будут химические и физические характеристики среды, тем меньше будет разнообразие видов, но зато выше численность особей, относящихся к одному и тому же виду. Такие взаимоотношения между числом видов и числом особей, с одной стороны, и степенью экстремальности условий - с другой, наблюдаются во многих экосистемах, например в горячих источниках, соленых озерах, кислых шахтных водах, в кишечнике и в сухих почвах. В экстремальных экосистемах преобладают те организмы, которые полностью приспособились к своему местообитанию и не растут, если соответствующий экстремальный фактор проявляется в меньшей степени. Это относится к крайне термофильным, психрофильным, галофильным, алкалифильным, осмофильным и другим «экстремофильным» микроорганизмам.
17.1.2 Водные экосистемы
Во многих экосистемах земной биосферы микроорганизмы занимают важное место или являются единственными формами жизни. Все экосистемы рассмотреть невозможно, поэтому мы должны ограничиться лишь несколькими. В эту небольшую выборку следовало бы включить почву как пример наземной экосистемы и океаны и моря как пример водных экосистем. Изучение плодородного пахотного слоя почвы в высшей степени интересно, но из-за наличия здесь огромного числа весьма разнородных организмов в очень малом пространстве экосистема почвы очень сложна. Поэтому мы сосредоточим свое внимание на водных системах, особенно в связи с тем, что большая часть микробиологических превращений происходит в водной среде. Типичными водными экосистемами являются океаны, моря, озера, пруды и проточные водоемы.
Океаны. Морская микробиология составляет часть биологии моря и как наука еще очень молода. Первичными продуцентами в море служат одноклеточные водоросли - фитопланктон. В пищевую цепь входят бактерии, простейшие, членистоногие и рыбы. Хотя океаны поглощают и накапливают наибольшее количество солнечной энергии, они участвуют в продукции пищи очень слабо; лишь 5-10% производимого на Земле белка образуется в океане. Продуктивность распределена здесь очень неравномерно. Эту неравномерность первичной и вторичной продукции органического вещества можно проиллюстрировать данными обуловах рыбы. Открытый океан, занимающий 90% водной поверхности нашей планеты, дает только 0,7% всего улова рыбы; на прибрежные зоны, составляющие около 10% площади, приходится 54% улова, а естественные области подъема глубинных вод (0,1% поверхности) дают 44% общего улова. Улов рыбы, очевидно, тесно связан с общей продукцией биомассы. Его распределение позволяет ясно увидеть зависимость первичного образования биомассы от количества питательных веществ, главным образом нитратов и фосфатов. Поэтому поступление богатых питательными веществами сточных вод в мировой океан не приводит к его загрязнению - более того, оно служит предпосылкой для производства биомассы в морях. Без постоянного притока таких веществ даже моря не могут давать больших уловов.
Интересные с точки зрения бактериологии превращения происходят в краевых участках моря, около устьев рек (в эстуариях), в засоленных маршах и в области солоноватых вод. Повсеместное присутствие в морской воде сульфата приводит к тому, что в анаэробных зонах и микроместообитаниях благодаря деятельности сульфатредуцирующих бактерий образуется сероводород, который оказывает воздействие на все остальное бактериальное сообщество.
Галофильные бактерии из прибрежных зон морей пока не настолько хорошо изучены, как они того заслуживают. Не только для познавательных целей, но и для решения практических задач необходимо постоянно держать в поле зрения и морские бактерии. В настоящее время большое внимание уделяется микроорганизмам сточных вод и вопросам распада трудноразложимых веществ в таких водах. Сточные воды загрязнены не только органическими примесями, но и значительными количествами солей, в том числе сульфатов. Таким образом, в них создаются условия, сходные с условиями морских экосистем. Эти соображения относительно биологии сточных вод подчеркивают необходимость уделять больше внимания микробиологическим превращениям, происходящим в экосистемах морей.
Озера. Наука об озерах и прудах (лимнология) позволила нам лучше понять частичные кругообороты и их интеграцию. Озера и более мелкие пресные водоемы представляют собой хорошо отграниченные, легко поддающиеся описанию водные экосистемы. В них имеются как аэробные, так и анаэробные зоны. Такие зоны можно обнаружить и в большинстве почв; но если в почве они сосредоточены поблизости друг от друга в очень тесном пространстве и потому их трудно изучать, в озерах такие зоны весьма обширны и легко поддаются исследованию. Однако есть основания полагать, что результаты лимнологических исследований в принципе можно перенести и на почву с ее микрогетерогенностью.
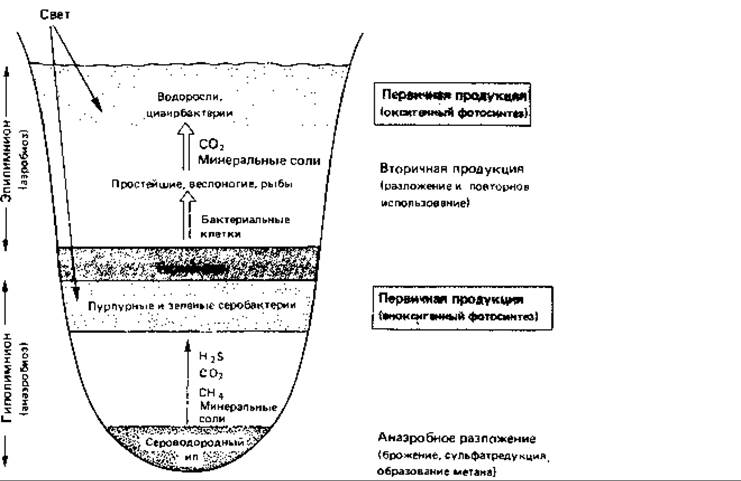
Рис. 17.1. Схематический вертикальный разрез эвтрофного озера - пример водной экосистемы. Слой температурного скачка (термоклин, или хемоклин) отделяет аэробную область от анаэробной. В обеих областях осуществляется первичное образование органического вещества путем фотосинтеза. Анаэробные условия начинают создаваться в результате анаэробного распада в донных осадках.
На биологические процессы в озерах и прудах большое влияние оказывают физические свойства воды. Вода имеет наибольшую плотность при 4°С. С увеличением глубины меняется температура воды; может наблюдаться более или менее устойчивая слоистость (стратификация) в зависимости от времени года (рис. 17.1). Стратификация характерна для озер двух типов. К одному типу относятся пресноводные озера умеренных широт. Весной холодная вода озера прогревается солнцем; поверхностный слой воды становится теплым, его плотность уменьшается. Этот слой называют эпилимнионом. Он лежит поверх более холодного слоя воды - гиполимниона. Слои разделены переходной зоной, называемой термоклином или металимнионом; эта граница между слоями иногда бывает очень резкой. В глубоких озерах такое разделение может сохраняться в течение всего лета. В результате процессов аэробного распада растворенный в воде кислород, начиная с придонного слоя, расходуется, и в гиполимнионе создаются анаэробные условия. Эпилимнион соприкасается с кислородом воздуха, перемешивается с помощью ветра, и поэтому, как правило, условия здесь остаются аэробными. Это приводит к образованию градиентов окислительно-восстановительного потенциала и химических параметров в области термоклина, в связи с чем термоклин называют также хемоклином. Осенью происходит охлаждение эпилимниона. Если температура эпилимниона становится ниже температуры гиполимниона, происходит перемешивание обоих слоев, чему способствуют осенние бури. В случае полного перемешивания глубинные слои воды поднимаются вверх и вновь обогащаются кислородом. Благодаря этому ежегодно восстанавливается равномерное распределение питательных веществ, которыми богаты глубинные воды. Озера с полным перемешиванием воды называют голомиктическими. Зимой может происходить обратное расслоение. Температура воды на глубине составляет 4°С, а над ней находится слой более холодной воды, имеющей меньшую плотность, а иногда и слой льда. Когда весной температура в поверхностном слое поднимается выше 4°С, стратификация снова нарушается.
Если глубинные воды, богатые питательными веществами, попадают на поверхность, начинается массовое размножение цианобактерий и зеленых водорослей («цветение воды»). Масштабы превращений вещества и продукции биомассы зависят от количества пииательных веществ в водоеме: в богатых этими веществами (эвтрофных) озерах такие превращения весьма интенсивны, а в бедных (олиготрофных) едва заметны.
От голомиктических отличают меромиктические и амиктические озера, в которых происходит лишь частичное перемешивание или его не происходит вовсе, так что образуется стабильный анаэробный гиполимнион («монимолимнион»), сохраняющийся независимо от времени года. Такая перманентность стратификации свойственна главным образом тропическим озерам, где температура поверхностного слоя воды редко опускается ниже температуры глубинного слоя. Но меромиктические озера встречаются и в умеренных широтах. Стабильность стратификации в них чаще всего поддерживается высокой соленостью глубинной воды (в рукавах фиордов) или особыми географическими условиями.
На примере голомиктического озера можно описать биологические процессы, которые приводят к летнему расслоению и продолжаются несколько месяцев. В пронизанном лучами света эпилимнионе фитопланктон (диатомеи, жгутиковые, зеленые водоросли, цианобактерии) продуцирует биомассу. Обычно из окружающей среды в озеро поступает дополнительный органический материал. Часть этого органического вещества, в особенности частицы, содержащие целлюлозу, опускается на дно озера и разлагается. В начальной аэробной стадии разложения расходуется кислород, и на дне создаются анаэробные условия. В результате анаэробного распада образуются органические продукты брожения Н2, H2S, СН4 и СO2. Поскольку конвекции не происходит, эти продукты поступают из донных отложений в толщу воды очень медленно. Один только метан - главный продукт анаэробной цепи питания в донных осадках - выделяется в виде пузырьков газа. На своем пути к поверхности водоема часть метана переходит в раствор и окисляется использующими этот газ аэробными бактериями. Быстрое потребление кислорода в гиполимнионе обусловлено ускоренным распределением метана и ростом метанокисляющих бактерий. В конце концов во всем гиполимнионе создаются анаэробные условия.
Как только в гиполимнионе исчезает кислород, там снова начинаются превращения с участием анаэробных микроорганизмов. Первичные продукты брожения используются для восстановления нитрата и сульфата. Основная масса сероводорода образуется при восстановлении сульфата в толще воды (рис. 17.2). Гиполимнион и термоклин- истинный рай для анаэробных бактерий. При наличии H2S и достаточной интенсивности света ниже термоклина растут пурпурные и зеленые серобактерии, образующие второй слой с высокой первичной продукцией биомассы. В этой зоне можно найти формы, обладающие газовыми вакуолями, такие как Lamprocystis, Amoebobacter, Thiodictyon, Thiopedia, Pelodictyon и Ancalochloris, а также передвигающиеся с помощью жгутиков виды Chromatium и Thiospirillum. Образование биомассы за счет аноксигенного фотосинтеза значительно; об этом свидетельствует количество инфузорий, веслоногих и ветвистоусых рачков, которые живут непосредственно над термоклином и питаются там фототрофными бактериями. Образуемый серными пурпурными бактериями сульфат быстро снова восстанавливается до H2S; при этом продукты, выделяемые фототрофными бактериями, вероятно, служат для сульфатредуцирующих бактерий донорами водорода.
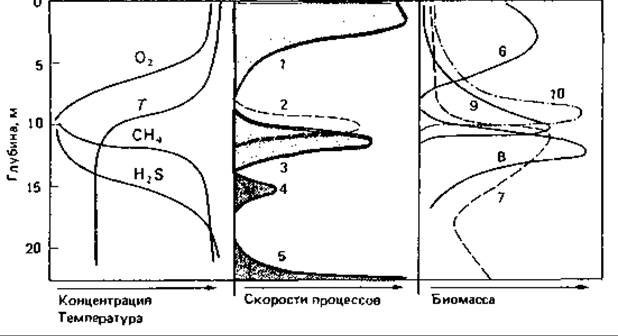
Рис. 17.2. Модельное представление вертикального профиля пресноводного озера умеренных широт с указанием концентраций, скоростей кругооборота и биомасс. В основу этой схемы положены данные Ю И. Сорокина (1970), В. М. Горленко, Г. А. Дубининой и С. И. Кузнецова (1977), а также Дж. Овербека (1972). T - температура, °С; 1 - фиксация СO2 на свету путем оксигенного фотосинтеза; 2 - фиксация СО2 в темноте; 3 - фиксация СO2 на свету путем аноксигенного фотосинтеза; 4 и 5 - сульфатредукция; 6-10-биомасса (мг/мл); 6 - водоросли и цианобактерии; 7 - общая бактериальная масса; 8 - фототрофные бактерии; 9 - простейшие; 10 - ветвистоусые и веслоногие рачки.
Для зоны термоклина тоже характерна высокая биологическая активность. Здесь развиваются некоторые цианобактерии, способные переносить присутствие сероводорода и отсутствие O2, в том числе Oscillatoria limnetica.
Описанные взаимоотношения представлены на рис. 17.1 и 17.2. Из этих схем ясно видно, что в стратифицированном озере имеются два типа водной среды, в которых идет первичное образование биомассы за счет фотосинтеза: в слоях, близких к поверхности эпилимниона, происходит оксигенный фотосинтез, a в верхнем слое гиполимниона - аноксигенный фотосинтез.
Проточные водоемы. В естественных, незагрязненных проточных водоемах часто бывает так мало одноклеточных организмов, что вода кажется кристально прозрачной. Следует, однако, вспомнить, что суспензия, содержащая 106 бактерий в 1 мл, остается на вид незамутненной. До тех пор пока загрязнение водоемов было незначительным, участок ручья или реки длиной в несколько километров мог минерализовать весь легко разлагаемый органический материал, поступающий из прибрежных селений. Состав микрофлоры и микрофауны в проточном водоеме служит хорошим индикатором степени его загрязнения. Если в водоеме еще встречаются дафнии - значит, вода чистая. Присутствие «гриба сточных вод» Sphaerotilus natans указывает на сильное загрязнение органическими веществами, а запах сероводорода свидетельствует об анаэробной сульфатредукции, т. е. служит сигналом тревоги.
Очистка сточных вод. Очистное сооружение в принципе представляет собой проточный водоем, в котором при участии грибов и бактерий (аэробных и анаэробных) происходит разложение органических веществ. Загрязнения в сточных водах могут быть различного рода в зависимости от того, что сбрасывается, - только фекалии и бытовые отходы или также навоз, сточные воды боен или другие промышленные отходы. Во многих случаях сточные воды содержат тяжелые металлы или устойчивые органические соединения. Цель очистки сточных вод состоит в освобождении их от твердых и жидких минеральных и органических веществ, прежде чем эти воды попадут в ручьи и реки. Особые усилия требуются для миурализации органического материала микробиологическим путем.
Содержание органических веществ, разлагаемых микробами, оценивают по так называемому «биологическому потреблению кислорода» (ВПК). Это количество кислорода, необходимое микроорганизмам для окисления органического материала в процессе дыхания. Например, ВПК 5 - это количество кислорода (мг), которое будет потреблено микроорганизмами в процессе разложения органических веществ за 5 дней. «Химическое потребление кислорода» (ХПК) означает количество кислорода, необходимое для полного химического окисления тех же веществ до СO2 и Н2O.
Для очистки сточных вод в очистных системах используются различные технические приемы, однако при этом осуществляются в принципе одни и те же основные этапы:
1) удаление относительно легко осаждаемых твердых частиц в пескоуловителе и в первичном отстойнике;
2) микробиологическое окисление растворенных органических веществ с применением активного ила либо с использованием биофильтра;
3) инкубация осадка, удаленного из первичного и вторичного отстойников, в анаэробных условиях в метантенке, где в результате образуется метан и выпадает осадок. После обезвоживания из этого осадка можно получать компост и использовать его в качестве удобрения или сжигать (рис. 17.3).
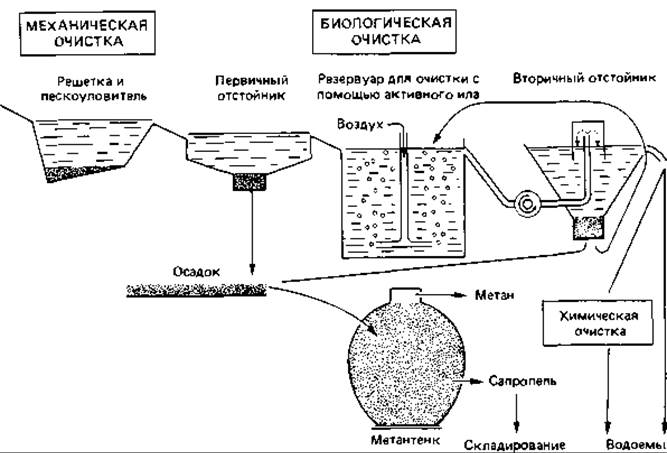
Рис. 17.3. Схема сооружения для механической и биологической очистки сточных вод.
Затем очищенная, осветленная вода сбрасывается в реки - непосредственно или через водоприемник. Эта вода еще содержит продукты минерализации - ионы фосфата, нитрата, аммония и другие. В результате ее сброса в реке может создаться такой избыток питательных веществ, что это вызовет увеличение первичной продукции. Для того чтобы избежать такой эвтрофизации водоемов, можно либо использовать очищенные сточные воды для орошения полей или удобрения лесных почв, либо добавить к обычной процедуре еще один этап очистки и путем денитрификации освобождать сточные воды хотя бы от связанного азота. Дополнительно их можно очищать путем химического осветления, а именно осаждения ионов фосфата с помощью солей железа. Возможно проведение и других мероприятий по очистке сточных вод.