Общая микробиология - Шлегель Г. 1987
Прокариоты. Систематический обзор
Метанобразующие бактерии и другие архебактерии (группа 13)
К группе 13 в определителе Берги отнесены метанобразующие бактерии. По многим причинам их в последнее время объединяют с двумя другими группами под общим названием «архебактерии». От всех остальных бактерий - эубактерий - они отличаются рядом важных признаков, и их находят в местообитаниях с экстремальными условиями. Эти условия сходны с теми, которые, вероятно, существовали на заре развития земной жизни. Среди архебактерий есть литоавтотрофы и гетеротрофы, анаэробы и аэробы. Все они имеют много общих признаков, свойственных только им. Однако между отдельными представителями архебактерий, по-видимому, имеются такие же различия в отношении формы клетки, ее компонентов и обмена веществ, какие существуют и между представителями эубактерий. Современное состояние исследований позволяет выделить среди архебактерий три группы: метанобразующие бактерии, галобактерии и термоацидофильные бактерии.
Общие признаки. У архебактерий есть целый ряд общих особенностей. Это касается состава клеточной стенки, липидов, аппарата транскрипции и трансляции, простетических групп и коферментов, механизма автотрофной фиксации СО2, а также способа получения энергии. Хотя результаты исследований еще весьма фрагментарны, мы попытаемся сделать общий обзор этой группы.
Клеточная стенка не имеет пептидогликанового остова, в лучшем случае она содержит псевдомуреин или только белки и полисахариды. Этим объясняется нечувствительность архебактерий к антибиотикам, нарушающим синтез клеточных стенок у эубактерий, - к пенициллину, цефалоспорину и D-циклосерину.
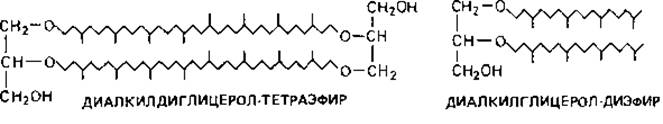
Плазматическая мембрана вместо глицериновых эфиров жирных кислот содержит глицериновые эфиры с С20 (фитанил)- и С40(бифитанил) - изопреноидными алкилами. В качестве нейтральных липидов встречаются также свободные С15- и С30-изопреноидные углеводороды:
Архебактерии отличаются от эубактерий по составу ДНК-зависимых РНК-полимераз; эти ферменты состоят у них из более чем четырех субъединиц и нечувствительны к антибиотикам рифампицину и стрептолидигину. Рибосомные нуклеиновые кислоты (16S и 5S) существенно отличаются по последовательности нуклеотидов. Трансляция нечувствительна к хлорамфениколу, однако тормозится дифтерийным токсином, который не действует на эубактерии, но подавляет синтез белка у эукариот.
Среди коферментов и простетических групп встречаются компоненты, которые хотя и сходны с таковыми у эубактерий и эукариот, но не идентичны им: производное 5-деазарибофлавина F420, никельтетрапиррольный фактор F430, метаноптерин, кофермент М и другие. Эти факторы были обнаружены у метанобразующих бактерий (разд. 9.4). Однако обобщающие выводы делать еще рано.
Автотрофная фиксация СО2 осуществляется не с помощью рибулозо- бисфосфатного цикла. Метаногенные бактерии используют для этого путь ацетил-СоА (разд. 9.4). Такой путь, однако, функционирует и у ряда эубактерий.
Процесс преобразования энергии (регенерации АТР), вероятно, основан на создании протонного потенциала и функционировании АТР-синтетазы, и его можно рассматривать как примитивное «анаэробное дыхание». Акцептором электронов служит СО2, сера или (у немногих архебактерий) молекулярный кислород.
Метанобразующие бактерии. Среди бактерий, образующих метан, встречаются практически все формы, известные у эубактерий: кокки (Methanococcus vannielii), палочки (Methanobacterium formicicum), короткие палочки (Methanobrevibacter ruminantium, М. arboriphilicus), спириллы (Methanospirillum hungatei), пакеты кокков (Methanosarcina barkeri) или нити (Methanothrix soehngenii) и даже формы, образующие пластины из клеток (Methanoplanus łimicola). Существуют мезофильные и термофильные виды (Methanobacterium thermoautotrophicum, Methanothermus fervidus). В настоящее время уже выделяют шесть семейств, а число описанных родов и видов быстро растет. Содержание GC варьирует в пределах от 27 до 61 мол. %. Об экологии и метаболизме этих бактерий будет сказано в разд. 9.4.
Галобактерии. Роды Halobacterium и Halococcus1содержат экстремально галофильные формы. Это аэробы и гетеротрофы. Их находят в солеварнях, где добывают морскую соль. Во время массового размножения галобактерий, содержащих каротиноиды, вода кажется ярко-красной. Они лучше всего растут в 3,5-5 М растворе NaCl. Еще одна их особенность - это способность использовать в своем метаболизме энергию света (разд. 12.3).
Термоацидофильные бактерии. В этой группе до сих пор объединяют неметаногенные термофильные архебактерии, имеющие между собой мало общего. Среди них есть автотрофы и гетеротрофы, крайне ацидофильные и нейтрофильные, аэробные и анаэробные представители.
Для Sulfolobus acidocaldarius местом обитания служат кислые горячие источники, где эта бактерия окисляет серу до сульфата (разд. 11.2). Особняком стоит Thermoplasma acidophilum; так же как и микоплазма, она не имеет клеточной стенки. Эта бактерия лучше всего растет при 59°С и pH 1-2. Место ее обитания-самонагревающиеся терриконы угольных шахт, но она была найдена и в горячих источниках. Ее геном - наименьший из всех изученных геномов непаразитических бактерий (1∙109). Она способна расти в дрожжевом экстракте как аэробный гетеротроф. Перечисленным аэробным видам противостоит группа анаэробных видов, объединяемых под названием Thermoproteales. Они были выделены из горячих источников на склонах вулканов и на дне морей. Это крайние термофилы (оптимумы от 85 до 105°С), и им свойствен тип метаболизма, называемый «серное дыхание» (разд. 9.3); они окисляют Н2 и восстанавливают элементарную серу до сероводорода. Среди них встречаются факультативные автотрофы (Thermoproteus tenax), облигатные автотрофы (Thermoproteus neutrophilus, Pyrodictium occultum) и гетеротрофные бактерии (Desulfurococcus, Thermococcus, Thermodiscus).
1 В последнее время описаны три новых вида галобактерий: Haloarcula, Natrobacterium и Natrococcus. - Прим. ред.
По мере изучения этой группы организмов становится все яснее, что архебактерии, судя по особенностям их клеточных компонентов, очень рано отделились от эубактерий; при этом большинство из них - вероятно, прямые потомки тех прабактерий, которые научились использовать доступные на заре эволюции неорганические Н-доноры (Н2) и Н-акцепторы (СО2, серу) и содействовали появлению уже более 3 млрд, лет назад отложений восстановленного углерода в осадочных породах.