Общая микробиология - Шлегель Г. 1987
Вирусы: распространение и структура
Отношение вирусов и плазмид к образованию опухолей (онкогенезу)
Возникновение злокачественных (раковых) опухолей может иметь различные причины, однако во всех случаях к этому причастен генетический материал клетки - ее ДНК. Что бы ни привело к образованию опухоли (раковому перерождению), последующим ростом ткани управляет ДНК безудержно делящихся опухолевых клеток. В основе превращения нормальной клетки в злокачественную - опухолевой трансформации — лежит перенос или иное изменение ДНК. Агент, вызывающий пролиферацию клеток,-это продукт гена. До сих пор, правда, не удается создать общую теорию, которая охватывала бы все формы ракового перерождения, однако изучение злокачественных опухолей, вызванных вирусами и плазмидами, уже сейчас позволяет сделать далеко идущие выводы.
Мы рассмотрим три примера онкогенеза: 1) образование опухолей у растений, 2) развитие опухолей у животных под действием ДНК-вирусов и 3) развитие опухолей у животных под воздействием РНК-вирусов (ретровирусов).

Рис. 4.15. Опухоль на стебле Kalanchoe blossfeldiana, образовавшаяся в результате прививки Agrobacterium tumefaciens в кору растения. (Фото U. Kaiser.)
Образование опухолей у растений. У многих растений встречаются опухоли корневой шейки. Эти разрастания ткани уменьшают поток питательных веществ между подземными и надземными частями. У многих растений такие опухоли можно вызвать экспериментально; типичные результаты получаются больше чем у половины изученных видов (рис. 4.15). Возбудителем является Agrobacterium tumefaciens - грам-отрицательная почвенная бактерия с перитрихальными жгутиками, сходная с представителями рода Rhizobium. Бактерии проникают в ткань через поврежденные участки и размножаются в межклетниках. Бывают вирулентные и авирулентные штаммы A. tumefaciens; вирулентные содержат большую плазмиду, так называемую Ті-плазмиду. После заражения ткани плазмиды проникают в растительные клетки.
Плазмидная ДНК прочно интегрируется в хромосомную ДНК растительных клеток и вызывает их опухолевый рост. Путем прививки таких клеток можно передать опухоль здоровому растению; таким образом, после того как клетки претерпели опухолевую трансформацию, бактерия и ее плазмида становятся уже ненужными. Интегрированная ДНК плазмиды ответственна также за способность клеток вырабатывать новые ферменты, с помощью которых синтезируются аминокислоты октопин и нопалин, так называемые опины. Эти аминокислоты могут использоваться бактерией A. tumefaciens в качестве источника углерода и азота. Благодаря Ті-плазмиде Agrobacterium получает, таким образом, преимущественный доступ к продуктам фотосинтеза растения: Ті-плазмида обеспечивает образование аминокислот, которые могут быть усвоены только этой бактерией.
Наряду с этим Ті-плазмида представляет собой естественный генный вектор для переноса чужеродной ДНК (см. разд. 15.3, рис. 15.21) в растения. Гены, определяющие опухолевый рост, можно выделить из плазмиды и заменить другими генами. Из тканей, состоящих из клеток, трансформированных видоизмененной плазмидой, удавалось регенерировать целые растения табака, которые росли совершенно нормально и вдобавок ко всему синтезировали опины. Таким образом, гены чужеродной ДНК передавались как доминантные факторы в соответствии с обычными законами наследственности.
Онкогенез, вызываемый у животных ДНК-вирусами. Исследования канцерогенеза у животных нередко проводятся на культурах тканей. Если перенести клетки животных, например из органов кур или хомячков, или фибробласты человека в подходящую питательную среду, то на внутренней стенке культурального сосуда они начнут размножаться. Обычно клетки продолжают расти лишь до тех пор, пока не начнут соприкасаться между собой. Из-за контактного торможения образуется только однослойный клеточный газон. Если же эти нормальные клетки инфицировать опухолеродным вирусом, то контактное торможение снимается, клетки продолжают размножаться и начинают надвигаться друг на друга. Многослойный рост наблюдается только у клеток, претерпевших опухолевую трансформацию. Из клеточной массы легко выделить отдельные клетки и таким путем получить чистые линии (клоны) трансформированных клеток.
Вирусы полиомы и SV40 («Обезьяний вирус 40») относятся к группе паповавирусов. Они содержат двухцепочечные кольцевые молекулы ДНК. В эксперименте вирус можно перенести для размножения в клетки тканевой культуры. Размножаясь в некоторых (так называемых пер- миссивных) клетках, вирус вызывает их лизис, и по мере его размножения клетки гибнут. В других (непермиссивных) клетках вирус ведет себя иначе. В этом случае размножение вируса подавляется, и примерно в одной из 105 клеток вирусная ДНК интегрируется в клеточную ДНК. Такое включение вирусной ДНК в геном клетки-хозяина может приводить к опухолевой трансформации. В трансформированной клетке образуется белок (Т-антиген), который запускает репликацию клеточной ДНК, и в результате начинается размножение клеток. Инъекция такого рода трансформированных клеток животным приводит к быстрому образованию опухолей.
Онкогенез, вызываемый у животных РНК-вирусами. К образованию опухолей у животных могут быть причастны также и РНК-вирусы-ретровирусы. Они относятся к икосаэдрическим вирусам с оболочкой и содержат (+)РНК-геном (одноцепочечную РНК). В качестве онкогенных вирусов они, например, вызывают саркому Рауса у кур и лейкемию у мышей. Название «ретровирусы» связано с тем, что в их размножении участвует обратная транскриптаза (см. конец раздела 15.1). РНК этих вирусов не может воспроизводиться путем простой репликации - необходима ее предварительная транскрипция в ДНК с последующей интеграцией этой ДНК в одну из хромосом клетки-хозяина. Интеграция - необходимый этап репродукции вируса; только интегрированная вирусная ДНК будет транскрибироваться. Так как интеграция в клеточную ДНК входит в жизненный цикл вируса, частота интеграции очень велика. Вероятно, вирусная ДНК может включаться в клеточную в любом месте.
Размножение вируса не приводит к лизису клетки. Нуклеокапсид образуется внутри клетки, перемещается затем к плазматической мембране и выходит наружу, одетый в оболочку из этой мембраны. Интегрированная ДНК ретровируса реплицируется вместе с геномом клетки- хозяина и поэтому содержится в каждой клетке опухоли (саркомы). Опухолевый рост клеток обусловлен экспрессией вирусного гена «src». Этот ген кодирует белок, который, по-видимому, представляет собой киназу, фосфорилирующую белки. Можно думать, что эта киназа участвует в преобразовании дифференцированной клетки в клетку эмбрионального типа.
Недавно была выяснена последовательность оснований в вирусной РНК. Оказалось, что она сходна с последовательностью одного из генов человека. Отсюда можно заключить, что src - это ген животного происхождения, который в результате неточной транскрипции был включен в РНК вируса вместе с вирусными генами и закрепился в ней. Это мог быть ген, кодирующий важный для эмбриональной клетки фактор роста. Таким образом, опухоли, вызываемые ретровирусами, в конечном счете обусловлены переносом какого-то гена животного происхождения в клетку животного.
Вироиды. Недавно удалось установить, что возбудителями некоторых опухолей растений являются маленькие «голые», т.е. лишенные белковой оболочки, свободные молекулы РНК. Их назвали вироидами. Это замкнутые в кольцо одноцепочечные молекулы с длиной цепи примерно в 360 нуклеотидов (мол. масса 12 ∙104 Да).
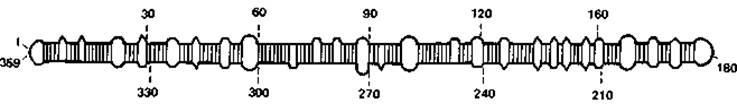
Таким образом, они в десять раз меньше инфекционных РНК самых мелких из известных до сих пор вирусов, и, следовательно, это самые мелкие возбудители болезней. Вироиды вызывают болезни картофеля, цитрусовых, огурцов, хризантем, хмеля, кокосовых пальм и других растений.