Общая микробиология - Шлегель Г. 1987
Клетка и ее структура
Эукариотическая клетка (эуцит)
Первые сведения об общей организации и тонкой структуре клетки были получены с помощью оптических методов. По мере совершенствования оптических приборов и улучшения техники микроскопирования росли и наши знания о микроморфологии клетки и ее отдельных компонентов. Дальнейшее развитие световой микроскопии, а также ультрафиолетовой и электронной микроскопии позволило существенно повысить разрешающую способность оптических приборов; темнопольная и фазово-контрастная микроскопия облегчила наблюдение живой клетки. Несомненно, и поныне микроскопия, особенно электронная, в сочетании со сложной предварительной обработкой биологического материала остается важнейшим методом исследования. Для получения дополнительных сведений о молекулярном уровне приходится прибегать к косвенным физическим и химическим методам, с помощью которых стало возможным выделять и исследовать отдельные клеточные компоненты.
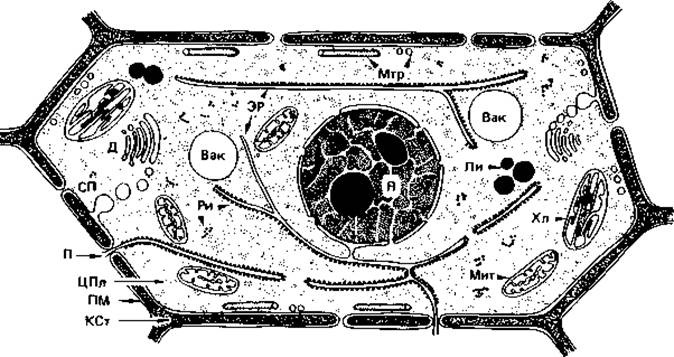
Рис. 2.1. Комбинированная схема строения эукариотической (растительной) клетки (по Зитте). Вак - вакуоли; Д - диктиосомы; КСт - клеточная стенка; Ли - липидные капельки; Мит - митохондрии; Мтр- микротрубочки; П - поры с плазмодесмами; ПМ-плазматическая мембрана; СП - секреторные пузырьки (экзоцитоз); Хл - хлором ласты; ЦПл - цитоплазма; Я - ядро.
Из разрушенных клеток (гомогенатов) методом дифференциального центрифугирования были выделены отдельные органеллы и фракции, что позволило изучать их биохимическими методами. Благодаря комбинации оптических и биохимических методов удалось быстро выяснить структуру и функции органелл и иных составных частей клетки. В результате этих исследований было установлено, что эукариоты и прокариоты по многим особенностям отличаются друг от друга.
Всякая клетка состоит из цитоплазмы и ядерного материала, снаружи ее ограничивает плазматическая мембрана. Этот протопласт может быть окружен еще клеточной стенкой, выполняющей в основном механические функции; стенка имеется у растительных клеток и у клеток большинства бактерий.
Мы кратко рассмотрим основные особенности эукариотических и прокариотических клеток (эуцитов и протоцитов). Типичным представителем эуцитов может служить эмбриональная растительная клетка (рис. 2.1).
Клеточное ядро. Структура ядра и способ его деления - важнейшие и самые характерные признаки, отличающие эукариотическую клетку (рис. 2.2) от прокариотической. Ядро (интерфазное) окружено ядерной оболочкой - двуслойной перфорированной мембраной. ДНК, несущая генетическую информацию, распределена между отдельными субъединицами-хромосомами, которые становятся видимыми только во время деления ядра. Ядро делится путем митоза (рис. 2.2); митоз обеспечивает 1) идентичную редупликацию генетического материала (что видимым образом проявляется в продольном расщеплении хромосом и удвоении их числа) и 2) передачу полного набора хромосом каждому из дочерних ядер. Как происходит удвоение хромосом, еще не вполне выяснено. Распределение хромосом может быть прослежено с помощью светового микроскопа и поэтому известно уже давно. В то время как интерфазное ядро в световом микроскопе кажется лишенным какой-либо четкой структуры, во время его деления хромосомы укорачиваются и становятся видимыми. Затем они располагаются в одной плоскости, образуя экваториальную пластинку. В результате сокращения нитей веретена половинки продольно расщепившихся хромосом расходятся; веретено исчезает, хромосомы становятся невидимыми и дочерние ядра окружаются снова ядерными оболочками.
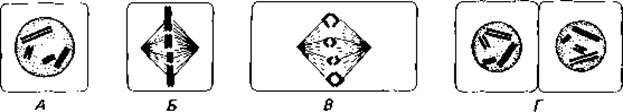
Рис. 2.2. Схема митотического деления диплоидной клетки. Ядро содержит по две хромосомы каждого вида, полученные от родителей (на схеме-красные и черные). В результате митоза хромосомы распределяются поровну между обеими дочерними клетками. А. В профазе становятся видимыми уже продольно расщепившиеся хромосомы; ядерная оболочка исчезает. Б. В метафазе хромосомы располагаются в экваториальной плоскости. В. В анафазе половинки расщепившихся хромосом оттягиваются нитями веретена к противоположным полюсам. Г. В телофазе дочерние хромосомы, вновь продольно расщепившиеся, окружаются ядерной мембраной, после чего ядро переходит в интерфазное состояние.
У всех высших растений и животных в процессе полового размножения происходит смена ядерных фаз. При оплодотворении половые клетки (гаметы) и их ядра сливаются, образуя зиготу. Отцовское и материнское ядра вносят при оплодотворении одинаковое число хромосом (n); таким образом, ядро зиготы содержит двойной хромосомный набор (2n). Иными словами, гаметы - гаплоидные клетки (т. е. клетки с одним набором хромосом), а соматические клетки - диплоидные (с двумя наборами). Поэтому при образовании гамет следующего поколения число хромосом в клетке (2n) должно уменьшиться вдвое (2n/2 = n). Совокупность процессов, приводящих к уменьшению числа хромосом, называют мейозом или редукционным делением (рис. 2.3). Мейоз - важнейший процесс у организмов, размножающихся половым путем: он приводит к двум результатам: 1) к перекомбинированию отцовских и материнских наследственных факторов (генов) и 2) к уменьшению числа хромосом. Мейоз начинается с конъюгации хромосом-каждая хромосома соединяется с соответствующей (гомологичной) хромосомой, происходящей от другого родителя. Во время конъюгации путем разрыва и перекрестного воссоединения (кроссинговера) может происходить обмен фрагментами одинаковой длины между гомологичными хромосомами. Затем следует двукратное разделение спаренных расщепившихся хромосом, и в результате образуются четыре клетки, каждая из которых имеет гаплоидное ядро. Таким образом, в процессе мейоза не только происходит «перетасовка» хромосом материнского и отцовского происхождения, но может произойти и обмен сегментами между гомологичными хромосомами. Оба процесса приводят к новым сочетаниям генов (к их рекомбинации).
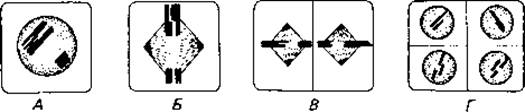
Рис. 2.3. Схема мейоза. Мейоз приводит к перекомбинированию отцовских и материнских генов и к уменьшению вдвое числа хромосом. Сначала происходит конъюгация гомологичных хромосом (А), и гомологи обмениваются участками (кроссинговер); в результате двукратного образования веретен (Б и В) хромосомы расходятся; при этом расходятся также и гомологичные хромосомы (В); в результате получаются четыре клетки с гаплоидными ядрами (Г).
У многих низших растений, включая водоросли, а также у простейших редукция числа хромосом происходит сразу после образования зиготы, так что организм оказывается гаплоидным. У растений с гетеро- фазной (антитетической) сменой поколений (мхов, папоротников) гаплоидные поколения чередуются с диплоидными.
Хромосомы эукариот состоят из нитей ДНК, с которыми связаны многочисленные белки. Часть этих белков составляют гистоны - основные белки. ДНК и гистоны, по-видимому, ассоциированы друг с другом весьма упорядоченным образом и образуют нуклеосомы — структурные субъединицы хромосом.
В ядре на хромосомной ДНК образуется мРНК. Через поры в ядерной мембране она транспортируется в цитоплазму. В интерфазном ядре можно видеть ядрышко; оно содержит ядрышковую ДНК, в которой заключена информация для построения рибосомной РНК и, вероятно, транспортной РНК. Обе эти РНК синтезируются в ядрышке и тоже переходят в цитоплазму. В эмбриональных клетках и в яйцеклетках содержится по нескольку или помногу ядрышек.
В эукариотической клетке ядро - важнейший, но не единственный носитель генетической информации. Часть такой информации содержится в ДНК митохондрий и хлоропластов.
Цитоплазма. Протопласт снаружи окружен плазматической мембраной. Для эукариотической клетки характерно выраженное подразделение цитоплазмы на множество -обособленных пространств. Эта компартментализация отчасти создается в результате впячивания плазматической мембраны с образованием цистерн и пузырьков; однако помимо этого в цитоплазме эуцитов содержатся митохондрии и (у растений) хлоропласта, которые со всех сторон окружены мембранами.
Из впячиваний плазматической мембраны образуется эндоплазматический ретикулум (ЭР). Часть ЭР образует наружную ядерную мембрану и, таким образом, окружает ядро; в ядерной оболочке имеются поры, которые обеспечивают беспрепятственный транспорт нуклеиновых кислот, белков и метаболитов между ядерным пространством и цитоплазмой. Часть мембран усеяна мельчайшими гранулами-рибосомами; это так называемый «шероховатый», или гранулярный, ЭР. На рибосомах осуществляется синтез белков. Свободно взвешенные в цитоплазме или прилегающие к ЭР рибосомы относятся к типу 80S.
Особой мембранной органеллой животных клеток является аппарат Гольджи. Сходные органеллы растительных клеток называют диктиосомами. Они состоят из пакетов уплощенных мембранных пузырьков, так называемых цистерн. Аппарат Гольджи и диктиосомы обеспечивают секрецию различных продуктов, главным образом ферментов. Ферменты синтезируются на цистернах и накапливаются в них. Со временем такой пузырек отделяется, перемещается к плазматической мембране, сливается с ней и изливает при этом свое содержимое наружу. Этот процесс получил название экзоцитоза.
Митохондрии и хлоропласта. В эукариотических клетках есть еще два вида органелл, окруженных мембранами, - митохондрии и хлоропласты. Митохондрии осуществляют дыхание; эти образования изменчивой формы, богатые липидами, имеют две мембраны - наружную и сильно складчатую внутреннюю (с кристами или трубочками). Внутренняя мембрана содержит компоненты электрон-транспортной цепи и ATP-синтазу. В клетках водорослей и высших растений наряду с митохондриями имеются также хлоропласты. Внутренние мембраны хлоропластов (тилакоиды) содержат фотосинтетические пигменты и компоненты фотосинтетического транспорта электронов.
Эндоцитоз. Для эукариотической клетки характерна способность поглощать пищу в виде растворенных веществ или оформленных твердых частиц. Общеизвестным примером поглощения твердых частиц служит фагоцитоз - захватывание их лейкоцитами крови или амебами. Если речь идет о поглощении жидких питательных веществ, говорят о пиноцитозе. Оба способа поглощения внеклеточного материала объединяют под названием эндоцитоза.
Способность эукариот захватывать оформленные твердые частички, в том числе и живые клетки, имеет фундаментальное биологическое значение. В эндоцитозе можно усмотреть предпосылки для возникновения эндосимбиоза и его механизм. Обычно твердые частички, поглощенные путем фагоцитоза амебой, перевариваются ею и полностью лизируются. В ряде случаев, однако, результатом может быть внутриклеточный симбиоз. Наиболее известный пример такого эндосимбиоза-ассоциация клеток бобовых растений с бактериями рода Rhizobium в корневых клубеньках (разд. 13.1). Подобного рода эндосимбионты широко распространены у эукариот (разд. 17.2.1). Способность эукариотических клеток приобретать эндосимбионтов говорит в пользу теории о симбиотическом происхождении хлоропластов и митохондрий. Прокариоты к эндоцитозу неспособны.
Эндосимбиотическая гипотеза. Клеточные органеллы эукариот имеют много фундаментальных общих черт с прокариотическими клетками. Они содержат кольцевые молекулы ДНК, их рибосомы относятся к типу 70S, а мембраны содержат компоненты электрон-транспортной цепи (флавины, хиноны, Fe-S-содержащие белки, цитохромы) и выполняют функцию дыхательного или фотосинтетического преобразования энергии. Согласно симбиотической гипотезе, митохондрии происходят от бесцветных аэробных бактерий, а хлоропласты - от цианобактерий, сделавшихся эндосимбионтами каких-то примитивных эукариотических клеток. В дальнейшем должна была произойти очень большая специализация; функция регенерации АТР была передана клеточным органеллам. Наружная мембрана эукариотической клетки не содержит компонентов электрон-транспортной цепи. С другой стороны, клеточные органеллы тоже не самостоятельны; они, правда, обладают собственными молекулами ДНК, однако значительная часть информации, необходимой для синтеза их белков, находится в клеточном ядре. Примером может служить рибулозобисфосфат-карбоксилаза - ключевой фермент автотрофной фиксации СO2 у зеленых растений. Она состоит из 8 больших и 8 малых субъединиц. Информация для построения больших субъединиц содержится в ДНК самих хлоропластов, а для синтеза малых - в ядре. По этой причине невозможно обеспечить развитие органелл вне клетки, так что гипотеза об эндосимбиозе недоступна для прямой проверки.
Двигательные органеллы. У всех эукариотических клеток, имеющих жгутики или реснички (у простейших, водорослей, сперматозоидов и клеток мерцательного эпителия), строение их одинаково. На поперечном срезе видно, что на периферии жгутика располагаются девять двойных нитей, а в центре - две одиночные нити (структура «9 + 2»), Снаружи вся эта система одета плазматической мембраной. Основание жгутика закреплено в наружном слое цитоплазмы при помощи базального тельца, или блефаропласта. Блефаропласт представляет собой производное самовоспроизводящейся органеллы (центриоли).