ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
ОСНОВИ ПОРІВНЯЛЬНОЇ ЕМБРІОЛОГІЇ
Ембріологія (гр. embrion - зародок) - наука про будову та розвиток зародка (ембріогенез). В поняття ембріогенезу включають період розвитку зародка від моменту запліднення до народження, або вилуплення з яйця, або закінчення метаморфозу. Сучасна ембріологія як наука і навчальна дисципліна дещо ширше трактує це поняття, включаючи сюди будову та розвиток статевих клітин (прогенез) і ранній період розвитку організму після народження.
Ембріогенез відбувається у декілька стадій з поступовими якісними і кількісними змінами. Розрізняють наступні періоди ембріогенезу: прогенез, запліднення, дроблення з утворенням бластули, гаструляція і диференціювання зародкових листків з утворенням зачатків тканин (гістогенез), органів (органогенез) та систем органів (системогенез) плоду.
ПРОГЕНЕЗ
Зрілі статеві клітини на відміну від соматичних містять гаплоїдний набір хромосом. Всі хромосоми гамети, за винятком одної статевої, називаються аутосомами. У чоловічих гаметах ссавців містяться статеві хромосоми Х або Y, а в жіночих - лише Х-хромосома.
Яким же чином відбувається зменшення удвічі числа хромосом, адже всі клітини тіла - диплоїдні і лише статеві - гаплоїдні?
Цей спосіб поділу клітин, внаслідок якого відбувається редукція числа хромосом і перехід клітин із диплоїдного стану у гаплоїдний, носить назву мейоз (гр. meiosis - зменшення).
Мейоз
Мейоз складається із двох послідовних поділів, в процесі яких редуплікація ДНК відбувається тільки один раз - перед першим поділом.
Особливістю першого поділу мейозу є складна і сильно розтягнута у часі профаза І, в якій розрізняють п’ять стадій: лептотена, зиготена, пахитена, диплотена, діакінез (Рис. 31).
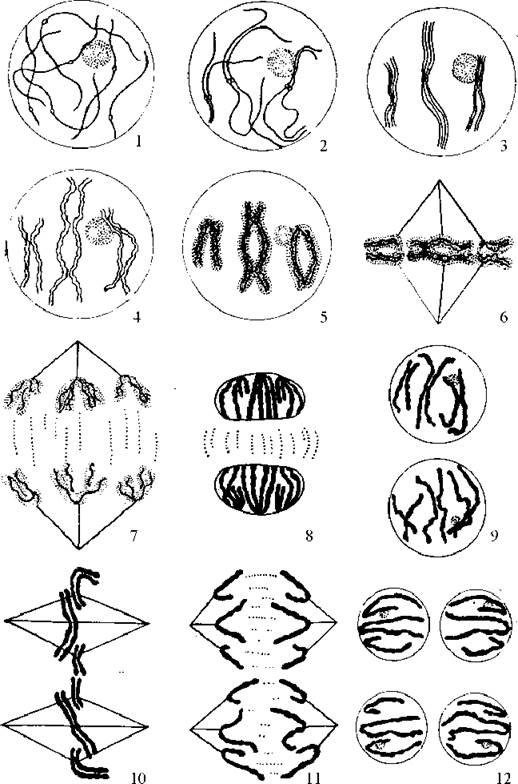
Рис. 31. Послідовні стадії мейозу:
1 - лептотена, 2 - зиготена, 3 - пахитена, 4 - диплотена, 5 - діакінез, 6 - метафаза І, 7 - анафаза І, 8 - телофаза І, 9 - інтерфаза, 10 - метафаза ІІ, 11 - анафаза ІІ, 12 - телофаза ІІ.
Лептотена (гр. leptos - тонкий) - стадія тонких ниток, нагадує ранню профазу мітозу. Починається конденсація хромосом, кожна хромосома складається з двох хроматид, з’єднаних центромерою.
Зиготена (гр. zygoo - з’єднання) - стадія ниток, що зливаються. Гомологічні хромосоми кон’югують, і всі гомологи об’єднуються у біваленти. Гомологи розпізнають один одного завдяки наявності в їхніх ДНК специфічних для кожної пари гомологів ділянок - z-ДНК.
Пахитена (гр. pachys - товстий) - стадія товстих ниток. Хромосоми потовщуються внаслідок спіралізації. Основна подія цієї стадії - кросинговер - обмін ділянками гомологічних хромосом. Електронна мікроскопія виявляє між кон’югатами в місцях їх щільного прилягання наявність білкових структур складної будови. Їх називають синаптонемальними комплексами, вважається, що саме вони забезпечують кросинговер.
Диплотена - стадія подвійних ниток - починається відштовхуванням гомологів і появою чітких хіазм (структура схожа на грецьку букву «хі» - Х). Стає видно, що кожен гомолог складається з двох хроматид. У складі біваленту чітко розрізняються 4 хроматиди. Такий бівалент називають тетрадою. Синаптонемальні комплекси зберігаються лише у місцях хіазм, тут триває кросинговер. У хроматидах з’являються ділянки розкручування, де синтезується РНК.
Диплотена - найбільш тривала стадія профази І (у людини - від 10-12 до 50 років). У цей період відбувається комплементарний синтез м-РНК, що сильно відрізняє мейоз від мітозу, де синтез РНК припиняється на початку профази. Цю стадію ще називають стадією «лампових щіток». Хромосоми розпушуються і утворюють петлі, які своїм виглядом нагадують колишні лампові щітки (Рис. 32). Чому під час мейозу дозволений синтез РНК? В цей час клітина росте і накопичує запаси хімічних компонентів апарату трансляції, необхідні для ранніх стадій розвитку зародка.
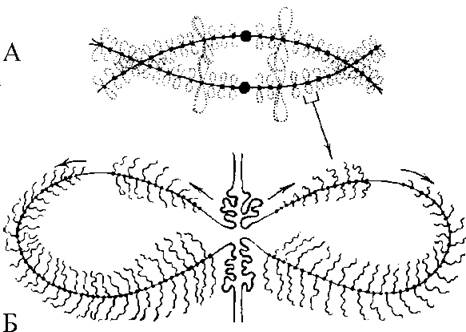
Рис. 32. Схема диплотенних хромосом, стадія «лампових щіток»:
А - ділянка біваленту з двома хіазмами, парне розташування бічних петель; Б - пара петель на сестринських хроматидах. Показано два цистрони на кожній петлі.
Діакінез - стадія відокремлювання подвійних ниток. Біваленти стають більш компактними і містять лише по дві термінальні хіазми. Руйнується ядерна мембрана, зникає ядерце, формується мітотичне веретено. Ця стадія є перехідною до метафази І, коли біваленти розташуються в екваторіальній площині клітини.
Анафаза І, як і під час мітозу, супроводжується розходженням хромосом до полюсів клітини. Але на відміну від мітозу до полюсів розходяться не сестринські хроматиди, а гомологічні хромосоми, які складаються з сестринських хроматид. Це призводить до того, що по різних клітинах розходяться алельні гени, розташовані у різних гомологах. Розподіл же ж гомологів по клітинах є зовсім випадковим. Редукції числа хромосом ще не відбулося, тому що кожен з наборів містить по 2n число хроматид. Але відбулась редукція генетичної різнорідності, тому що тепер у кожному з хромосомних наборів відсутні алельні гени.
Після телофази І може спостерігатись коротка інтерфаза без S- періоду (т.т. без редуплікації ДНК), і клітини починають наступний поділ, який не відрізняється від класичного мітозу: парні сестринські хроматиди проходять профазу і метафазу, в анафазі вони роз’єднуються і потім розходяться до дочірніх клітин, які містять вже по гаплоїдному набору хромосом.
Таким чином, мейоз є редукційним поділом: перший поділ є редукційним у загальногенетичному аспекті (втрачається генетична різнорідність), а другий - у морфологічному (вдвічі зменшується хромосомний набір). Після двох послідовних поділів з однієї диплоїдної клітини утворюється чотири гаплоїдних, кожна з яких відрізняється за своєю генетичною конституцією.