ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
ОСНОВИ ПОРІВНЯЛЬНОЇ ЕМБРІОЛОГІЇ
ПРОГЕНЕЗ
Статеві клітини
Чоловічі статеві клітини - сперматозоїди (у всіх хребетних рухаються), або спермії (якщо нерухомі) - розвиваються у дуже великій кількості: у людини об’єм сім’яної рідини, що виділяється під час еякуляції становить в нормі близько 3 мл. У ній міститься 3-3,5 · 108 сперматозоїдів. Вважається, що для того, щоби відбулось нормальне запліднення, сім’яна рідина повинна містити не менше 150 млн сперматозоїдів, при цьому концентрація сперматозоїдів в еякуляті повинна бути не меншою 6 · 107 мл-1. У деяких тварин кількість сперматозоїдів може сягати кількох мільярдів (риби, птахи, коні). У статевих шляхах жінки кількість сперматозоїдів після коїтусу становить близько 2 · 108, але лійки маткової труби, де відбувається зустріч з яйцеклітиною, досягають лише близько 200 сперматозоїдів.
За своїми розмірами в більшості випадків сперматозоїди - невеликі клітини, однак варіюють у широких межах (від 20 мкм у крокодила до 5000 мкм у тритона), при цьому, як бачимо, їхні розміри не залежать від розмірів тварини. Розмір сперматозоїдів людини сягає 70 мкм, вони здатні до активних рухів (швидкість руху становить 30-50 мкм/с) за допомогою биття джгутика. Приблизно через півгодини після еякуляції вони опиняються у порожнині матки, а через 1,5-2 години - в ампулярній частині маткової труби.
Елементарно в сперматозоїда розрізняють головку, шийку і хвіст (Рис. 33).
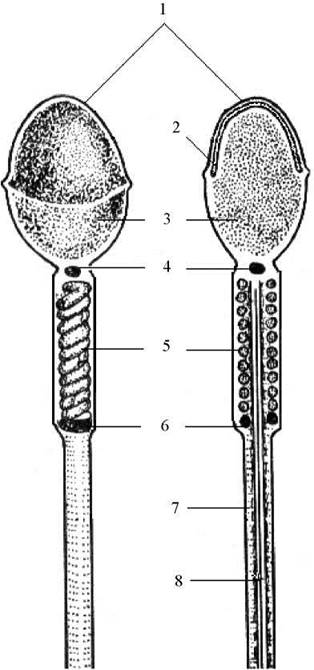
Рис. 33. Схема будови сперматозоїда:
1 - плазмолема чохлика, 2 - акросома, 3 - ядро, 4 - проксимальна центріоль, 5 - спіральні мітохондрії проміжної частини хвоста, 6 - частина дистальної центріолі, 7 - фібрилярний футляр головної частини хвоста, 8 - аксонема.
Основну частину головки займає досить ущільнене ядро, вкрите тонким шаром цитоплазми. У передній частині головки розташовується спеціальна органела сперматозоїда - акросома - яка містить набір ферментів, які у своїй сукупності прийнято називати спермолізинами (вони здатні руйнувати облолонки яйцеклітини).
Акросома є аналогом лізосоми і за своїм походженням є похідною комплексу Гольджі. Її мембрана досить щільно прилягає до плазмолеми переднього полюсу головки сперматозоїда, через що передня оболонка сперматозоїда при світловій мікроскопії здається дещо ущільненою і через це називається чохликом.
У шийці сперматозоїда розташовується проксимальна центріоль, а за нею дистальна, яка служить базальним тільцем для осьової нитки хвоста сперматозоїда - аксонеми. Досить часто частина дистальної центріолі, яка має вигляд кільця, розташовується каудальніше (лат. cauda - хвіст), на межі між проміжною і головною частинами хвоста.
У хвості виділяють проміжну, головну і термінальну (кінцеву) частини. На поперечному перерізі проміжної частини видно, що під плазмолемою розташовується мітохондріальна піхва у вигляді спірально закручених мітохондрій. Між нею і центрально розташованою аксонемою залягають окремі щільні фібрили білкової природи. Мітохондіальна піхва у головній частині відсутня, але замість окремих щільних фібри навколо аксонеми з’являється суцільний фібрилярний футляр. Термінальна частина хвоста містить окремі мікрофіламенти.
Тривалість життя і здатність до запліднення у сперматозоїдів різних видів тварин у певних оптимальних умовах значно різняться. У ссавців вони варіюють від декількох годин до декількох діб. У кислому середовищі або у присутності іонів двох- або трьохвалентних металів сперматозоїди аглютинують і втрачають здатність до запліднення.
Утворення чоловічих статевих клітин - сперматогенез - протікає в звивистих сім’яних канальцях сім’яників чоловіків. У сперматогенезі розрізняють чотири послідовні стадії: розмноження, ріст, дозрівання і формування (Рис. 34).
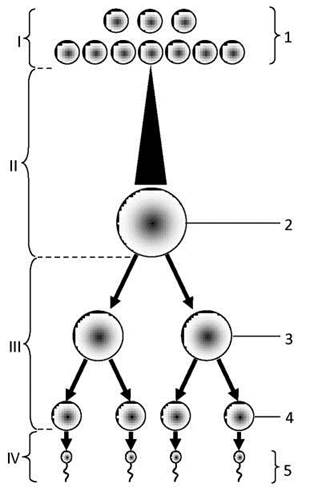
Рис. 34. Схема сперматогенезу:
І - період розмноження, ІІ - період росту, ІІІ - період дозрівання, ІV - період формування (сперміогенез);
1 - сперматогонії, 2 - сперматоцит 1-го порядку, 3 - сперматоцит 2-го порядку, 4 - сперматиди, 5 - сперматозоїди.
Початковою фазою сперматогенезу є розмноження сперматогоній - вихідних клітин, що займають найбільш периферичне (базальне) положення в сперматогенному епітелії (Рис. 35).
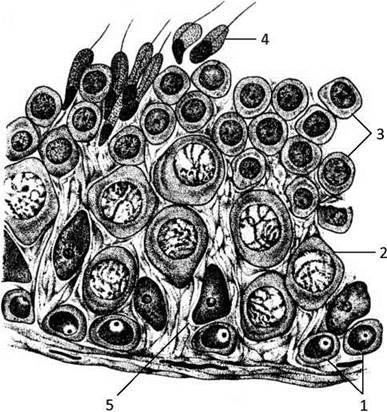
Рис. 35. Сперматогенний епітелій:
1 - сперматогонії, 2 - сперматоцит 1-го порядку, 3 - сперматиди, 4 - сперматозоїди, що формуються, 5 - клітини Сертолі.
У результаті повторних поділів стовбурних сперматогоній значна частина них відтискуються від базальної мембрани і, перестаючи поділятися, вступає у другу фазу - період росту, поступово перетворюючись у сперматоцити 1-го порядку. Інші ж сперматогонії продовжують поділятися, внаслідок чого запас сперматогоній у сім’яних канальцях не убуває, незважаючи на безупинне утворення сперматозоїдів. У періоді росту сперматоцити 1-го порядку значно збільшуються в об’ємі і готуються до редукційного поділу.
У періоді росту в зростаючих сперматоцитах 1-го порядку відбуваються ряд стадій профази I мейозу. Закінчується він на стадії диплотени утворенням тетрад. Сперматоцити 1-го порядку після цього вступають у період дозрівання, протягом якого кожен такий сперматоцит спочатку поділяється на два сперматоцити 2-го порядку. Вони відразу ж, не вступаючи в синтетичний період інтерфази, проходять другий поділ, результатом якого є утворення чотирьох сперматид.
Таким чином, кожна вихідна сперматогонія дає початок чотирьом сперматидам з гаплоїдним набором хромосом. Сперматиди більше не поділяються, але шляхом складної перебудови перетворюються в зрілі сперматозоїди. Ця трансформація складає четверту фазу сперматогенезу - період формування, або сперміогенезу.
Сперматиди являють собою невеликі округлі клітини з порівняно великими ядрами. Накопичуючись біля верхівок підтримуючих клітин - клітин Сертолі, сперматиди частково занурюються в їхню цитоплазму, що створює умови для формування сперматозоїдів зі сперматид.
У сперматидах біля ядра розташовуються пластинчастий комплекс, центросома і скупчення дрібних мітохондрій. Процес формування починається утворенням у зоні пластинчастого комплексу ущільненої гранули - акробласта, що прилягає до поверхні ядра. Надалі акробласт, збільшуючись у розмірах, у вигляді шапочки охоплює ядро, а в його середині диференціюється ущільнене тільце - акросома. Вона лежить у тім кінці сперматиди, що занурений у підтримуючу клітину. Центросома ж, що складається з двох центріолей, пересувається до протилежного кінця сперматиди. Проксимальна центріоль прилягає до поверхні ядра, а дистальна розділяється на дві частини. Від передньої частини дистальної центріолі починає формуватися аксонема хвоста сперматозоїда. Задня ж половина дистальної центриолі приймає вид колечка. Зміщаючись уздовж аксонеми, це колечко визначає задню границю проміжної частини хвоста сперматозоїда.
Ядро сперматиди поступово ущільнюється і товщає. Цитоплазма в міру росту хвоста сповзає з ядра і зосереджується в середній частині. Мітохондрії розташовуються по спіралі тіла сперматозоїда між проксимальної центріоллю і колечком. Цитоплазма сперматиди під час її перетворення в сперматозоїд сильно редукується, В області головки вона зберігається тільки у вигляді тонкого шару, що вкриває ядро та акросому; невелика кількість цитоплазми залишається в області шийки і, нарешті, вона дуже тонким шаром покриває аксонему хвоста.
В цілому процес сперматогенезу в людини триває близько 75 діб, але протікає протягом звивистого сім’яного канальця хвилеподібно. Тому на кожнім відрізку канальця всі клітини сперматогенного епітелію виявляються в одній з чотирьох фаз сперматогенезу.
Сперматогенний епітелій надзвичайно чутливий до ушкоджуючих впливів. При різних інтоксикаціях, у т.ч. алкогольних, авітамінозах, недостатності харчування й інших умов (особливо при впливі іонізуючими випромінюваннями) сперматогенез послаблюється або навіть припиняється, а сперматогенний епітелій атрофується. Аналогічні деструктивні процеси розвиваються при крипторхізмі (коли сім’яники не опускаються в мошонку, залишаючись у черевній порожнині), тривалому перебуванні організму в середовищіз високою температурою, пропасних станах і особливо після перев’язки чи перерізу сім’явивідних каналів.
Деструктивний процес при цьому уражає в першу чергу найбільш диференційовані яруси сперматогенного епітелію — сперматозоїди, що формуються, і сперматиди. Останні, набухаючи, нерідко зливаються в характерні округлі маси - так називані сім ’яні кулі, що плавають у просвіті канальця. Через те, що нижні шари сперматогенного епітелію (сперматогонії та сперматоцити 1-го порядку) при цьому зберігаються більш довгостроково, відновлення сперматогенезу після припинення дії ушкоджуючого агента іноді виявляється можливим.
Жіночі статеві клітини - овоцити або ооцити (лат. ovum - яйце) визрівають у значно менших кількостях, ніж сперматозоїди. Для деяких ссавців кількість яйцеклітин, які дозрівають за все життя, становить лише сотні штук. Разом з тим у деяких хребетних яйцеклітин буває значно більше (риби і амфібії).
Як правило, яйцеклітини нерухомі, мають кулясту форму і значно більший об’єм цитоплазми ніж сперматозоїди. Цитоплазма яйцеклітин містить специфічне включення - жовток (гр. lecithos - жовток). Залежно від кількості жовтка розміри овоцитів коливаються від кількох мікрометрів до кількох сантиметрів.
За кількістю жовтка розрізняють овоцити алецитальні (безжовткові), оліголецитальні (маложовткові) і полілецитальні, цитоплазма яких перевантажена жовтком. Кількість жовтка залежить від умов розвитку зародка і тривалості розвитку у зовнішньому середовищі.
Якщо жовток розподіляється по цитоплазмі рівномірно, як це спостерігається у оліголецитальних яйцеклітин, говорять про ізолецитальні (гр. isos - рівний) яйцеклітини.
У полілецитальних яйцеклітин жовток у більшою або меншою мірою зміщений до одного з полюсів клітини (вегетативного), а органели і ядро - до іншого (анімального). Такі яйцеклітин називаються телолецитальними (гр. thelos - кінець), а якщо жовток розташовується у центрі яйцеклітини - центролецитальними. Серед телолецитальних яйцеклітин розрізняють помірно телолецитальні, або мезолецитальні (як, наприклад, у амфібій), і різко телолецитальні (у плазунів і птахів).
Перехід до наземного існування привів до появи у плазунів і птахів більш складних за організацією яйцеклітин. Збільшились їхні розміри, вони стали різко телолецитальними. Наземний розвиток зародка викликав появу вторинних і третинних оболонок.
У плацентарних ссавців у зв’язку з внутрішнім розвитком зародка за рахунок материнського організму відпала необхідність запасання жовтка, і виникли удруге (в еволюційному аспекті) оліголецитальні яйцеклітини.
В цілому яйцеклітини мають звичайну для всіх клітин будову: плазмолема, цитоплазма, ядро (Рис. 36). Слід, однак, виділити деякі особливості будови яйцеклітин. Яйцеклітини часто полярні, що зумовлене розташуванням жовтка у цитоплазмі, можуть мати вторинні і третинні оболонки, їхня цитоплазма містить величезні запаси хімічних елементів апарату трансляції: рибосоми, різні види РНК. Під цитолемою розташовуються невеликі за розмірами (200-600 нм) так звані кортикальні гранули. Цитоплазма містить специфічний вид включень - жовток - у вигляді гранул або пластинок. У їх центрі зосереджений кристалоїдної структури фосфопротеїн - фосфовітин, пухку периферію утворює ліпопротеїн - ліповітелін.
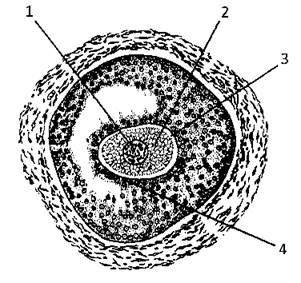
Рис. 36. Овоцит:
1 - ядро, 2 - цитоплазма, 3 - zona pellucida, 4 - corona radiata. Щоб відбулось запліднення, сперматозоїд має здолати променистий вінець.
В процесі росту і визрівання у яєчнику яйцеклітини оточуються спочатку одним, а потім декількома шарами плоских або кубічних клітин, що називаються фолікулярними (лат. foliculus - пухирець), які утворюють один із захисних бар’єрів овоцита - променистий вінець (corona radiata). За рахунок діяльності овоцита і фолікулярних клітин кругом першого утворюється зона, багата на глікозоаміноглікани (ГАГ) - прозора або блискуча зона - zona pellucida (ZP).
Зріла ZP містить густу сітку тонких ниток, що складаються з глікопротеїнів, і поділяється на два шари:
1) внутрішній, багатий на нейтральні ГАГ і
2) зовнішній, який містить переважно кислі ГАГ.
Глікозоаміноглікани - клас полісахаридів, побудованих з
дисахаридних одиниць, що періодично повторюються: одна з них є звичайною уроновою кислотою, друга - аміноцукор (N-ацетилглікозамін або N-ацетилгалактозамін). Якщо ГАГ сульфатовані, карбоксильовані або гідроксильовані, вони набувають кислих властивостей. Молекули ГАГ сильно гідрофільні. Зв’язані між собою молекули ГАГ утворюють гель, через який добре дифундують метаболіти. Розрізняють 5 типів ГАГ: гіалуронова кислота (нейтральна), хондроїтинсульфат, дерматансульфат, кератансульфат, гепарансульфат.
Основна маса зрілої ZP - глікопротеїни ZP: ZP1, ZP2, ZP3, з молекулярною масою відповідно 90-110, 64-76, 57-73 кД. Синтез білків ZP припиняється під час овуляції або одразу після неї.
ZP3 складається з поліпептиду (44 кД) і ланцюгів N- олігосахаридів та О-олігосахаридів. ZP містить приблизно 1012 молекул ZP3, які разом із ZP2 утворюють нитки довжиною 2-3 мкм і товщиною 7 нм. У складі ниток комплекс ZP2-ZP3 повторюється кожних 15 нм. Нерегулярним чином нитки з’єднані глікопротеїном ZP1, що призводить до утворення тримірної сітки, яка складає каркас прозорої оболонки (Рис. 37). В петлях цієї сітки розташовуються ГАГ.
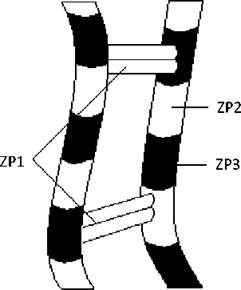
Рис. 37. Організація ZP-глікопротеїнів прозорої зони.
Прозора зона містить нитки, утворені молекулами глікопротеїнів ZP2 та ZP3. Глікопротеїн ZP1 нерегулярно з’єднує нитки між собою.
ZP3 є рецептором сперматозоїда, при цьому він суворо видоспецифічний. Інактивація або відщеплення О-олігосахариду від молекули ZP3 блокує зв’язування сперматозоїда з яйцеклітиною.
Сперматозоїд у плазмолемі головки містить рецептори до цього олігосахариду. Взаємодія ZP3 з рецептором на головці сперматозоїда дає старт акросомній реакції.
ZP2 є вторинним рецептором сперматозоїда. З початком акросомної реакції ZP2 додатково зв’язує сперматозоїд.
Під час запліднення відбувається хімічна модифікація ZP2 і ZP3, наслідком чого є блокада поліспермії.
Оогенез (овогенез). Оогенез (утворення яйцеклітини) протікає аналогічно сперматогенезу, але з деякими особливостями (Рис. 38).
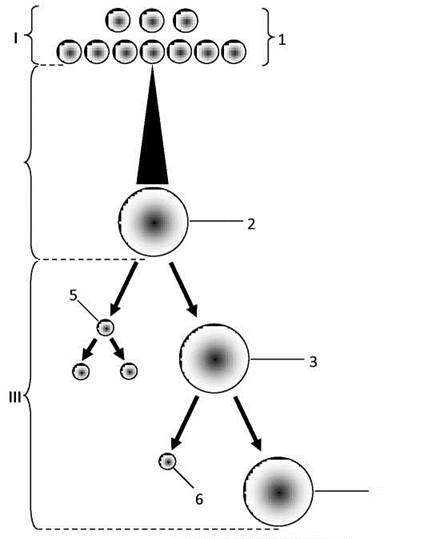
Рис. 38. Схема овогенезу:
І - період розмноження, ІІ - період росту, ІІІ - період дозрівання;
1 - оогонії, 2 - овоцит 1-го порядку, 3 - овоцит 2-го порядку, 4 - яйцеклітина, 5 - перше редукційне тільце, 6 — друге редукційне тільце.
Так, перша стадія - період розмноження - відбувається в період внутрішньоутробного розвитку та у перші місяці післянатального життя, коли в яєчнику зародка відбувається поділ оогоній і формування так названих примордіальних (лат. primordium - початок, виникнення) фолікулів.
Друга стадія - період росту - протікає у функціонуючому яєчнику і полягає у перетворенні оогонії примордіального фолікула через стадії первинного та вторинного фолікулів в овоцит 1-го порядку в третинному фолікулі (зрілий фолікул або графів пухирець). При цьому в ядрі зростаючого овоцита відбувається кон’югація хромосом і утворення тетрад, а в їхній цитоплазмі накопичуються жовткові включення.
Третя стадія - період дозрівання - відбувається в овоцитах 1-го порядку, що вийшли з яєчника в результаті овуляції. Період дозрівання, як і під час сперматогенезу, включає два поділи, причому другий наступає одразу за першим без інтерфази, що приводить до редукції числа хромосом вдвічі, і їхній набір стає гаплоїдним. При першому поділі дозрівання овоцит 1-го порядку поділяється на овоцит 2-го порядку і невелике редукційне тільце. Овоцит 2-го порядку одержує всю масу накопиченого жовтка і тому залишається настільки ж великим за об’ємом, як і овоцит 1-го порядку. Редукційне ж тільце являє собою дрібну клітину з невеликою кількістю цитоплазми, що одержує по одній діаді від кожної тетради ядра овоцита 1-го порядку.
При другому поділі дозрівання овоцит 2-го порядку поділяється на яйцеклітину і друге редукційне тільце. Перше редукційне тільце іноді теж поділяється на дві однакові дрібні клітини. У результаті цих нерівномірних розподілів з овоцита 1-го порядку утворюється одна яйцеклітина і три редукційних тельця. Четверта стадія - формування - в овогенезі відсутня.
Розвиток. Оогонія стає овоцитом 1-го порядку з того моменту, коли вона закінчує період розмноження і входить у період малого росту, що може довгостроково, але повільно продовжуватися в інфантильному віці і набуває швидкого плину після настання статевої зрілості. Повільно зростаючий овоцит оточується одним шаром плоских фолікулярних клітин, розташованих на базальній мембрані (Рис. 39). На початку великого росту фолікулярні клітини під впливом фолітропіну набувають призматичної форми, поділяються мітозами, і фолікулярний епітелій стає багатошаровим, одержуючи назву зернистої зони або зернистого шару. Навколо зростаючого овоцита відокремлюється щільна блискуча зона (zona pellucida).
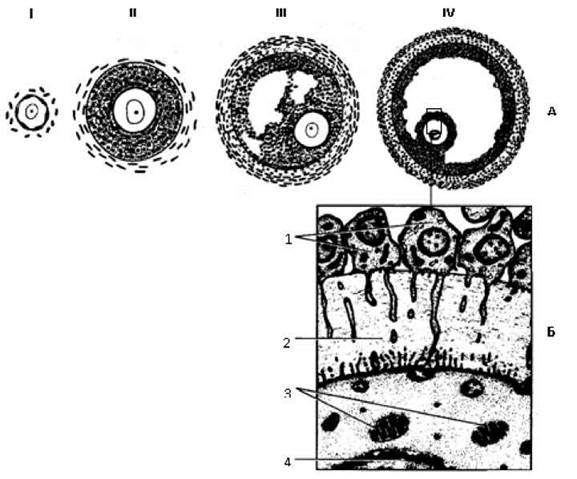
Рис. 39. Диференціювання фолікула.
А - стадії розвитку: І - примордіальний фолікул, II - первинний фолікул, III - вторинний фолікул, IV - третинний фолікул.
Б - зв’язок фолікулярних клітин з овоцитом: 1 - фолікулярні клітини, 2 - zona pellucida, 3 - мітохондрії, 4 - ядро.
В міру збільшення зростаючого фолікула його навколишня сполучна тканина ущільнюється, даючи початок зовнішній оболонці фолікула. У зовнішню оболонку вростають численні кровоносні капіляри і вона диференціюється на два шари: внутрішній, або судинний, і зовнішній, або фіброзний, утворений щільною сполучною тканиною. Утворюється так званий первинний фолікул.
Надалі клітини фолікулярного епітелію, що складають зернистий шар, посилено розмножуються і починають виробляти фолікулярну рідину, що спочатку накопичується між фолікулярними клітинами, а потім між фолікулярними клітинами утворюються порожнини. Так фолікул стає вторинним, що має діаметр близько 200 мкм. Один із вторинних фолікулів випереджає у рості інші та стає домінантним. Він швидко росте від 200 мкм до 1-2,5 см у діаметрі переважно за рахунок накопичення рідини у порожнинах, які по мірі росту зливаються в одну. Овоцит з його навколишнім шаром фолікулярних клітин, що називається променистим вінцем {corona radiata), відтискується до верхнього полюса зростаючого фолікула. Коли фолікул досягає максимуму свого розвитку, він одержує найменування третинного. Оскільки фолікул стає готовим до овуляції, він одержав додаткову назву передовуляторного. Ділянка зернистого шару, у якій залягає овоцит, називається яйценосним горбком.
Клітини променистого вінця, які безпосередньо оточують зростаючий овоцит, мають довгі гіллясті відростки, що проникають через блискучу зону, досягаючи поверхні овоцита. По цих відростках до овоцита від фолікулярних клітин надходять поживні речовини, з яких у цитоплазмі синтезуються складові жовтка.
Граафів пухирець досягає такого розміру, що випинає поверхню яєчника, причому яйценосний горбок з овоцитом виявляється у виступаючій частині пухирця. Подальше збільшення обсягу пухирця, переповненого фолікулярною рідиною, приведе до розтягування і витончення як його зовнішньої оболонки, так і білочної оболонки яєчника в місці прилягання цього пухирця.
Овуляція. Настання овуляції - розриву фолікула і викиду овоцита 1-го порядку в черевну порожнину - викликається дією лютеїнізуючого гормону (лютропін), коли виділення його гіпофізом різко збільшується. Механізм овуляції цілком ще не з’ясований. Овуляція пов’язана зі збільшенням припливу крові до капілярів внутрішньої оболонки і зростанням внутрішньофолікулярного тиску. Відому роль в овуляції може грати окситоцин. Перед настанням овуляції секреція окситоцина збільшується як рефлекс на подразнення нервових закінчень, що залягають у внутрішній оболонці, що обумовлюється підвищенням внутрішньофолікулярного тиску. Крім того, витонченню і розпушенню оболонки фолікула сприяють протеолітичні ферменти та гіалуронідаза.
Овоцит 1-го порядку, оточений фолікулярним епітелієм, з черевної порожнини потрапляє на фімбрії (ворсинки) лійки і далі в просвіт маткової труби. Тут швидко відбуваються поділи дозрівання, і утворюється зріла яйцеклітина, готова до запліднення.
Зіставлення сперматогенезу й овогенезу наведене в таблиці 1.
Табл. 1. Порівняльна характеристика гаметогенезу
Стадія |
||
Період |
сперматогенез і форми гоноцитів, які утворюються протягом нього |
овогенез і форми гоноцитів, які утворюються протягом нього |
Розмноження |
Відбувається у статевозрілому віці Сперматогонії |
Відбувається у внутрішньо-утробному періоді Оогонії (овогонії) |
Ріст |
Настає одразу за періодом розмноження Сперматоцити 1-го порядку |
Підрозділяється на: а) малий ріст б) великий ріст Ооцити (овоцити) 1-го порядку |
Дозрівання |
Поділ сперматоцитів рівномірний Сперматиди |
Поділ овоцитів нерівномірний Генетично однорідні яйцеклітина і редукційні тільця (Х) |
Формування |
Сперміогенез - трансформація сперматид у сперматозоїди, клітини генетично різнорідні (Х або Y) |
Відсутній |