ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
ОСНОВИ ПОРІВНЯЛЬНОЇ ЕМБРІОЛОГІЇ
ЗАПЛІДНЕННЯ
Заплідненням називають злиття чоловічої та жіночої статевих клітин, внаслідок чого відтворюється притаманний даному біологічному виду диплоїдний набір хромосом, різко зростає метаболізм і виникає якісно нова клітина - зигота (запліднена яйцеклітина або одноклітинний зародок). При цьому маса ядра збільшується удвічі, а об’єм цитоплазми через різницю об’єму яйця і сперматозоїда практично дорівнює об’єму яйцеклітини.
В процесі запліднення розрізняють три фази: 1) дистантна взаємодія і зближення гамет, 2) контактна взаємодія і активація яйцеклітини, 3) утворення чоловічого та жіночого пронуклеусів з наступним їх злиттям - сингамією.
Перша фаза - дистантна взаємодія і зближення гамет - спрямована на підвищення ймовірності зіткнення статевих клітин. Важлива роль при цьому належить хімічним речовинам, які на цій стадії виробляються овоцитами і сперматозоїдами. Так, у ссавців овоцит після овуляції, опинившись у фалопієвій трубі, починає продукувати речовину гіногамон І (ГГІ) (гр. gyne - женщина), яка стимулює рухливість всіх сперматозоїдів. Разом з тим кожний із сперматозоїдів виробляє речовину андрогамон І (АГІ) (гр. andrikos - чоловічий), який пригнічує рухливість інших, крім нього, сперматозоїдів.
Але ГГІ і АГІ лише активують або пригнічують рухливість сперматозоїдів. Виникає питання: а як сперматозоїди вибирають напрямок руху?
У статевих шляхах самки спостерігається пристінкова течія рідини із середини назовні. Сперматозоїдам притаманна властивість рухатись проти цієї течії - негативний реотаксис (гр. rheos - течія, taxis - напрямок).
Зрозуміло, що концентрація ГГІ у статевих шляхах має певний градієнт: ближче до овоциту вона зростає. Сперматозоїди рухаються за градієнтом концентрації ГГІ, т.т. спостерігається позитивний хемотаксис.
Вважається, що плазмолема яйцеклітини деполяризована таким чином, що на зовнішній її поверхні розташовуються катіони, а на поверхні сперматозоїда з початком акросомної реакції - аніони. Тому на близьких відстанях у дію вступає позитивний електротаксис - притягання різнойменних зарядів.
Таким чином, напрямок руху сперматозоїдів зумовлений негативним реотаксисом, позитивним хемотаксисом і позитивним електротаксисом, а швидкість руху - взаємодією ГГІ і АГІ.
Друга фаза - контактна взаємодія і активація яйцеклітини - вимагає від сперматозоїдів на відміну від попередньої - конкурентної - фази, спільних дій щодо руйнування оболонок яйцеклітини.
У безхребетних, риб, хвостатих амфібій, плазунів і птахів можлива поліспермія, коли у яйцеклітину одночасно проникають декілька сперматозоїдів, але у злитті ядер гамет, саме заплідненні, бере участь лише один. У ссавців у яйцеклітину проникає лише один сперматозоїд, таке явище називають моноспермією.
Виникає питання: навіщо тоді потрібні сотні мільйонів сперматозоїдів, що містяться в еякуляті?
На важкому і небезпечному для сперматозоїдів шляху до яйцеклітини значна частина їх в силу тих чи інших причин гине. Це так. Але, якщо пухку оболонку із фолікулярних клітин (променистий вінець) сперматозоїди проходять відносно легко, то прозора оболонка на їхньому шляху є досить важким для подолання бар’єром. Виявляється, що спермолізинів одного сперматозоїда не вистачає для руйнування ZP, а тому спільними діями сперматозоїди руйнують її. Як це відбувається?
Сперматозоїди, які пройшли променистий вінець із фолікулярних клітин, розштовхуючи їх, досягнувши прозорої зони, взаємодіють із ZPB-глікопротеїном, у відповідь на що у мембрані сперматозоїда відкриваються Na+- і Са2+-канали, що викликає каскад подій:
1) деполяризацію мембрани сперматозоїда, внаслідок чого її мембранний потенціал стає слабо позитивним;
2) підвищення у цитоплазмі сперматозоїда концентрації Ca2+, що активує Са2+-залежну фосфоліпазу, яка у свою чергу активує другі посередники - ц-АМФ і ц-АДФ, що приводить до
3) активації Н+-АТФази, що викликає підвищення внутрішньоклітинного рН, внаслідок чого відбувається фрагментація чохлика головки сперматозоїда (злиття плазмолеми сперматозоїда і мембрани акросоми (Рис. 40), через що спермолізини акросоми (трипсин, гіалуронідаза, акрозин, фосфатази, ліпази тощо) звільняються і починають руйнувати хімічні компоненти прозорої зони.
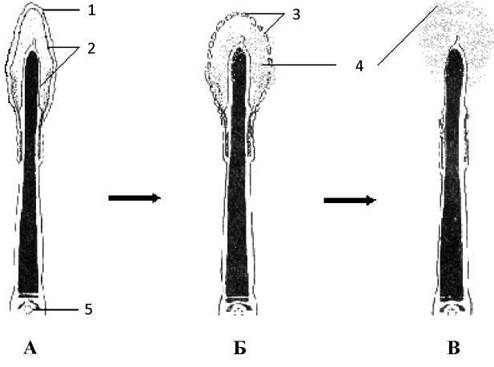
Рис. 40. Механізм акросомної реакції:
А, Б, В - послідовні стадії акросомної реакції.
1 - плазмолема, 2 - мембрана акросоми, 3 - мембранні пухирці, 4 - спермолізини, 5 - проксимальнацентріоль.
І як тільки у будь-якому місці поверхня овоцита оголюється, відбувається злиття (!) мембран сперматозоїда і яйцеклітини. У цитоплазму овоцита потрапляє вміст головки і проміжної частини хвоста сперматозоїда, а у плазмолемі овоцита деякий час зберігається вбудована у неї ділянка плазмолеми сперматозоїда (Рис. 41).
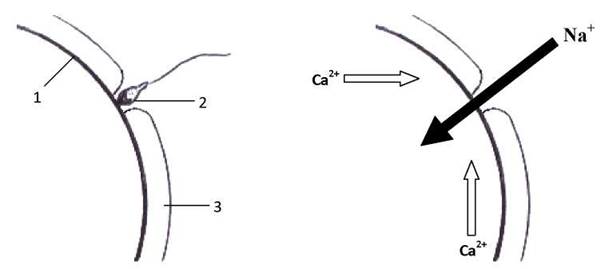
Рис. 41. Старт кортикальної реакції:
1 - плазмолема яйцеклітини, 2 - сперматозоїд, 3 - zonapellucida.
Саме вона ініціює початок так званої кортикальної реакції (Рис. 42):
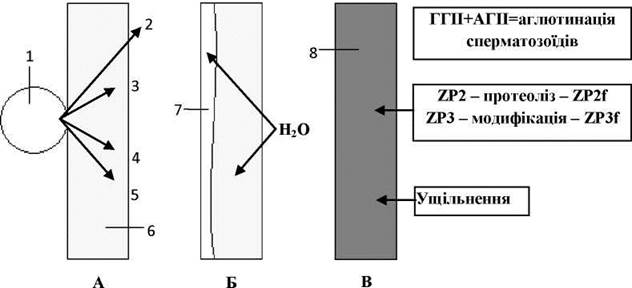
Рис. 42. Механізм кортикальної реакції:
А, Б, В - послідовні стадії кортикальної реакції.
1 - кортикальна гранула, 2 - ГГІІ, 3 - протеолітичні ферменти, 4 - гідрофільний фактор, 5 - фактор ущільнення, 6 - zona pellucida, 7 - перивітеліновий простір, 8 - оболонка запліднення.
1) через відкриті у плазмолемі сперматозоїда натрієві канали у цитоплазму яйцеклітини лавиноподібно прямують іони Na+, що викликає підвищення їх концентрації у кортикальному шарі цитоплазми;
2) за NA+/Са2+-обмінним механізмом у кортикальному шарі цитоплазми збільшується концентрація іонів Са2+;
3) підвищення концентрації іонів Са2+ викликає екзоцитоз кортикальних гранул, у яких містяться протеолітичні ферменти, гідрофільні та ущільнення фактори, а також молекули гіногамону ІІ (ГГІІ);
4) ГГІІ викидається за межі ZP, де викликає аглютинацію сперматозоїдів через високу спорідненість до вбудованого у їхню мембрану андрогамону ІІ (АГІІ);
5) протеолітичні ферменти розривають зв’язок ZP з плазмолемою яйцеклітини, утворюючи перивітеліновий простір, а також викликають хімічну модифікацію молекул ZP2- та ZP3- глікопротеїнів прозорої зони, через що сперматозоїди не можуть більше їх розпізнавати;
6) гідрофільний фактор викликає насичення ZP і перивітелінового простору водою;
7) фактор ущільнення ущільнює ZP, чим завершується утворення оболонки запліднення.
Вже через кілька хвилин після утворення оболонки запліднення у цитоплазмі яйцеклітини значно посилюються процеси метаболізму і синтезу білка. Разом з тим починається процес інтенсивного перемішування частин цитоплазми овоцита: утворюються зони
підвищеної концентрації органел, жовткових та пігментних гранул. Цей процес носить назву ооплазматичної сегрегації (лат. segregatio - відділення, відокремлення). Було встановлено, що вже на цьому етапі кожна ділянка цитоплазми овоцита певною мірою детермінована та у майбутньому дасть початок тій чи іншій структурі зародка. Такі ділянки отримали назву презумптивних (лат. praesumptio - припущення, основане на вірогідності) зачатків.
Третя фаза - утворення чоловічого та жіночого пронуклеусів з наступним їх злиттям - сингамією. Вміст головки сперматозоїда, опинившись у цитоплазмі яйцеклітини, повертається на 180°. Протягом перших 12 годин після проникнення сперматозоїда у яйцеклітину відбувається перебудова ядер і сперматозоїда, і яйцеклітини, вони набухають, з’являються ядерця - утворюються пронуклеуси. Останні наближаються один до одного, мігруючи до центру яйцеклітини. Відбувається редуплікація ДНК і дуплікація центріолей сперматозоїда. Диплосоми розходяться до полюсів яйцеклітини (яйцеклітина власних центріолей не має). Пронуклеуси вступають у мітоз. На стадії метафази хромосомні набори об’єднуються - утворюється синкаріон (лат. sin - зв’язок, гр. karion - ядро). Цей процес - сингамія - і є власне заплідненням, з’являється диплоїдна зигота. Внесок сперматозоїда у запліднення не вичерпується лише тим, що половина хромосом диплоїдної зиготи батьківські; частину генетичної інформації новий організм отримує разом з мітохондріями сперматозоїда. По ходу запліднення у яйцеклітині завершується мейоз, і детермінується генетична стать нового організму. Нарешті, сперматозоїд приносить з собою сигнальні білки дроблення (у зиготі виявляються два принесених сперматозоїдом білка з М.м. 14 і 18 кД, які містять одну й ту саму антигенну детермінанту. Антитіла до цього антигену блокують перші дроблення зиготи, не впливаючи на інші процеси).