ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
КЛІТИННІ МЕМБРАНИ
Основу клітинних мембран утворює елементарна біологічна мембрана. Вона являє собою ліпопротеїновий комплекс завтовшки 7-10 нм, до складу якого входять ліпіди (40 %), білки (60 %) і незначною кількістю вуглеводи.
Ліпіди утворюють бішар (Рис. 1, 2) і представлені фосфоліпідами, сфінгомієлінами і стероїдами (холестерин).
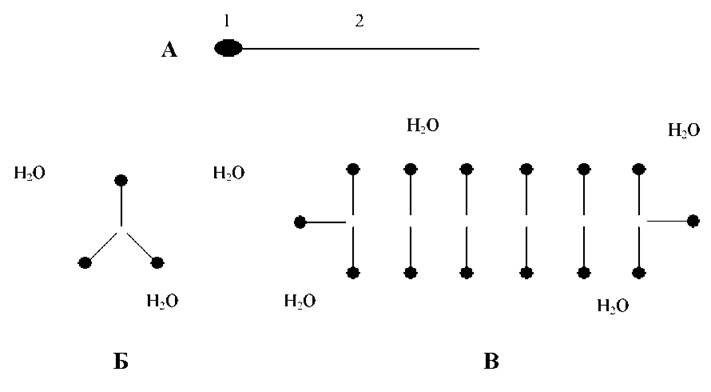
Рис. 1. Утворення ліпідного бішару.
А - просторова структура молекули ліпіду: 1 - головка, 2 - хвіст. Б - міцела. В - бішар.
Фосфоліпіди. Як і всі ліпіди, вони являють собою складні ефіри гліцерину та жирних кислот. Просторова структура молекули ліпіду представлена на Рис. 1. Як бачимо, вона має гідрофільну полярну головку і гідрофобний хвіст із жирної кислоти. Через це у водному розчині молекули ліпідів будуть розташовуватись відповідним чином: полярними головками до води.
Сфінголіпіди. Молекули сфінголіпідів містять луг з довгим ланцюгом (сфінгозин або аналогічну групу). Сфінголіпіди в значних кількостях містяться у оболонках клітин нейроглії і є основною складовою частиною мієліну.
Холестерин. Відноситься до класу стероїдів і крім участі у побудові мембран має надзвичайно важливе значення. По-перше, він є основою для синтезу стероїдних гормонів (статевих, глюко- і мінералокортикоїдів). По-друге, холестерин циркулює в організмі в складі двох фракцій ліпопротеїнів: ЛНЩ - ліпопротеїнів низької щільності та ЛВЩ - ліпопротеїнів високої щільності. Підвищення концентрації ЛНЩ викликає утворення на стінках судин атеросклеротичних бляшок - атеросклерозу. Якщо ж концентрація ЛНЩ перевищує концентрацію ЛВЩ більше ніж у п’ять разів, різко зростає ризик розвитку ішемічної хвороби серця. ЛВЩ, навпаки, видаляють холестерин із судин.
Білки - амінокислотні полімери. Більшість амінокислот мають полярні молекули, але є і неполярні (гліцин, аланін, валін, лейцин), тому білки у мембрані розташовуються теж відповідним чином: неполярними ділянками взаємодіють з хвостами ліпідів, полярними - з їхніми головками.
За місцем розташування у мембрані розрізняють білки інтегральні (трансмембранні, пронизують всю мембрану наскрізь), напівінтегральні (занурені у неї частково) і поверхневі (Рис. 2).
Функціонально серед них виділяють: структурні білки, які стабілізують структуру мембрани, рецепторні білки, канальні білки, що утворюють іонні канали, білки-ферменти і білки-переносники, які здійснюють переніс через мембрану іонів і дрібних молекул. Особливостей кожній мембрані надають білкові комплекси, більшість яких є рецепторними і ферментними системами.
Оболонка клітини (плазматична мембрана, плазмолема, цитолема)
Особливість клітинної оболонки полягає в тому, що вона не тільки оточує цитоплазму ззовні, але й забезпечує контакт клітини із зовнішнім середовищем, з усіма його шкідливими факторами. Тому до характерної для елементарної мембрани бар’єрної функції додається ще й захисна. Товщина плазмолема становить 20 нм, інколи значно більше (Рис. 2).
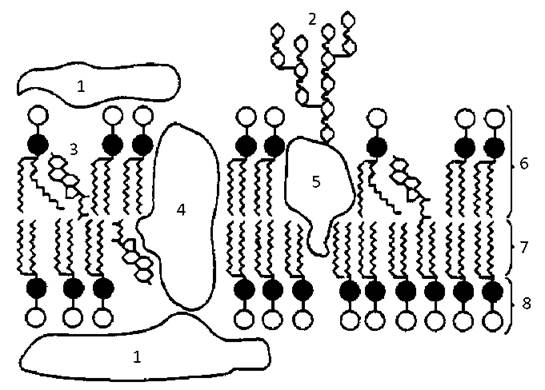
Рис. 2. Будова плазмолеми:
1 - поверхневі білки, 2 - олігосахарид у складі глікопротеїну, 3 - холестерин, 4 - інтегральний білок, 5 - напівінтегральний білок, 6 - шар фосфоліпідів, 7 - хвости жирних кислот, 8 - гідрофільні головки фосфорліпідів.
Ліпопротеїновий комплекс елементарної мембрани підстеляється ущільненим шаром цитоплазми, так званим кортикальним шаром, а ззовні вкритий шаром глікокаліксу. Останній утворений розгалуженими олігосахаридами, які, контактуючи з відповідними елементами мембрани, утворюють глікопротеїни і гліколіпіди. Між гілками олігосахаридів зустрічаються білки немембранного походження, наприклад, дипептидази в глікокаліксі епітелію кишечнику. Хімічний склад плазмолеми: білки - 60 %, ліпіди - 40 %, вуглеводи - 2-10 %.
Проникність плазмолеми визначається гідрофобним характером середини біліпідного шару.
Неполярні молекули вільно проникають через мембрану. Так, ендо- і екзоцитоз полярних молекул відбувається завжди з утворенням мембранних пухирців, а секреція чи поглинання неполярних речовин (наприклад, стероїдних гормонів) здійснюється без їхнього утворення. З цієї причини рецептори до стероїдних гормонів розташовуються усередині клітини (ядерні рецептори).
Полярні речовини не здатні проникати через мембрану. Саме з цієї причини рецептори до полярних молекул вбудовані в плазмолему, а передачу сигналу до інших клітинних структур здійснюють другі посередники (Рис. 3). Передачу сигналів від клітини до клітини здійснюють сигнальні молекули (перший посередник), які виробляються одними клітинами і специфічно впливають на інші - клітини-мішені. Специфічність впливу визначають присутні у клітинах-мішенях рецептори, що зв’язують тільки власні ліганди.

Рис. 3. Другий посередник - G-білок.
А - виключений стан, α-субодиниця зв’язана з ГДФ і не контактує з рецептором. Б - активований лігандом рецептор фосфорилює ГДФдо ГТФ, G-білок активується. В - після дисоціації активованого G-білка α-субодиниця зв’язується з ефектором і активує його. Г - α-субодиниця гідролізує ГТФ, інактивується і об’єднується зіншими субодиницями G-білка. Р - рецептор, Е - ефектор, Фн - фосфат.
G-білок складається з трьох субодиниць (СО) - α, β, γ. Вони розташовуються ближче до цитоплазматичної поверхні плазмолеми. У стані спокою всі СО об’єднані, α-СО асоційована з гуанозиндифосфатом (звідси абревіатура G-білок). Каскад подій, які розвиваються після взаємодії ліганда з рецептором, показаний на Рис. 3. Фосфорилювання ГДФ викликає дисоціацію α-СО, яка активує ефектор. Після дефосфорилювання ГТФ α-СО інактивується і займає своє місце у складі G-білка. Розрізняють чотири функціональні форми G-білка: Gs - активатор аденілатциклази, Gi - інгібітор аденілатциклази, Gp - активатор фосфоліпази С, Gt - активатор цГМФ-фосфодиестерази.
Рецепторами можуть бути білки, глікопротеїди і полісахариди, які вкривають всю поверхню клітини. Ліганди представляють найрізноманітніші групи біологічно активних речовин (гормони, медіатори, антигени тощо) і можуть бути навіть фізичними факторами (кванти світла).
Вибіркова проникність плазматичної мембрани, яка підтримує клітинний гомеостаз, оптимальний вміст в клітині води, іонів та ін., реалізується трьома шляхами: пасивним транспортом, полегшеною дифузією, активним транспортом (Рис. 4).
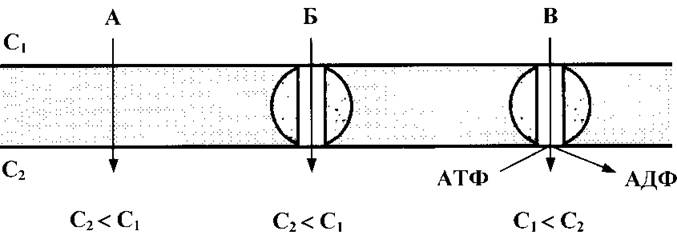
Рис. 4. Види транспорту речовин через плазмолему:
А - проста дифузія, Б - полегшена дифузія, В - активний транспорт. С1 та С2 - концентрації речовин відповідно зовні клітини і у цитоплазмі.
Пасивний транспорт - рух через плазмолему невеликих неполярних (О2, N2) і полярних молекул (СО2, Н2О) в обох напрямках за градієнтом концентрації або за електрохімічним градієнтом без витрат енергії.
Полегшена дифузія речовин відбувається за допомогою компонентів мембрани (канали і білки-переносники), в більшості випадків в одному напрямку (до клітини), за градієнтом концентрації і без витрат енергії.
За допомогою білків-переносників забезпечується транспорт глюкози, амінокислот, фосфатів, обмін Сl- на HCO3-, Na+ або К+ на Н+.
Іонні канали - іоноспецифічні мембранні пори, утворені декількома зв’язаними білковими субодиницями. Через канал за електрохімічним градієнтом проходять відповідні іони. Найбільш поширені іонні канали - для Na+, К+, Са2+, Сl-.
Активний транспорт - енергозалежний трансмембранний переніс іонів проти електрохімічного градієнту. Активний транспорт здійснюється інтегральними мембранними білками-ферментами - АТФазами. Найбільш поширені Na+,К+-, Н, К-, Са2-АТФази.
Плазматична мембрана забезпечує виконання ще двох функцій: ендоцитозу і екзоцитозу.
Ендоцитоз - поглинання клітиною речовин, частинок і мікроорганізмів (Рис. 5). Ендоцитоз здійснюється шляхом піноцитозу, фагоцитозу і опосередковуваного рецепторами ендоцитозу.

Рис. 5. Ендоцитоз (А) і екзоцитоз (Б):
1 - білки зливання, 2 - секреторний пухирець, 3 - інвагінація, 4 - ендосома.
Піноцитоз - процес поглинання клітиною рідини і розчинених речовин з утворенням невеликих пухирців.
Фагоцитоз - поглинання клітиною великих частинок (мікроорганізмів або залишків клітин). Фагоцитоз здійснюють спеціальні клітини - фагоцити (макрофаги, нейтрофіли). Фагосоми, що утворюються в процесі фагоцитозу, значно більші, ніж піноцитозні пухирці. Вони зливаються з лізосомами і утворюють фаголізосоми. На відміну від піноцитозу фагоцитоз індукується певними сигналами, що впливають на рецептори плазмолеми фагоцита. Подібними сигналами можуть служити антитіла, які оточують частинку, що фагоцитується.
Опосередковуваний рецепторами ендоцитоз характеризується поглинанням із міжклітинної рідини конкретних макромолекул. Мембранний рецептор розпізнає певну макромолекулу, утворюється комплекс ліганд-рецептор. Декілька (багато) таких комплексів концентруються в локальній ділянці плазмолеми, яка збоку цитоплазми облямована білком клатрином. Ця ділянка (облямована ямка) інвагінується, утворюється облямований клатрином пухирець. Після звільнення від клатрину утворюється ендосома, яка може зливатись з лізосомами.
Екзоцитоз - процес під час якого внутрішньоклітинні секреторні пухирці зливаються з плазмолемою, а їхній вміст звільняється із клітини. Як і під час ендоцитозу, важливу роль в цьому процесі відіграють спеціальні мембранні білки зливання.