ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
ЦИТОПЛАЗМАТИЧНІ ОРГАНЕЛИ І ВКЛЮЧЕННЯ
Цитоплазма містить органели, включення, фібрилярні компоненти цитоскелету. Її основу складає гіалоплазма (цитозоль).
Цитоплазматичні органели - це обов’язкові компоненти цитоплазми, які мають певну будову і виконують специфічні функції. В залежності від наявності мембрани, яка відмежовує вміст органели від цитозолю, розрізняють органели мембранні і немембранні (Рис. 6).
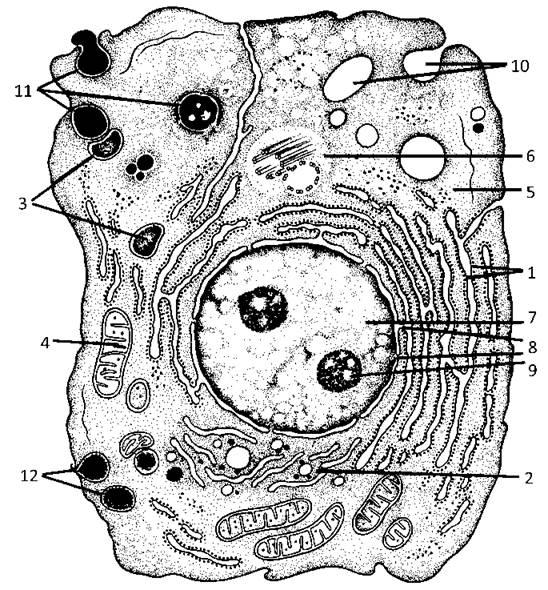
Рис. 6. Схема будови клітини:
1 - ГрЕПР, 2 - комплекс Гольджі, 3 - лізосоми, 4 -мітохондрія, 5 - рибосоми, 6 - центріолі, 7 - ядро клітини, 8 - ядерна оболонка, 9 - ядерце, 10 - піноцитоз, 11- фагоцитоз, 12 - секреція.
До мембранних органел відносять гранульований ендоплазматичний ретикулум, гладкий ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, мітохондрії та пероксисоми.
Немембранні органели представлені рибосомами, мікротрубочками, центріолями, війками та джгутиками, іншими фібрилярними структурами.
В залежності від функціонального навантаження розрізняють органели загального призначення і спеціальні.
Гранульований ендоплазматичний ретикулум (ГрЕПР) (К.Р. Портер, 1945) являє собою систему плоских замкнених мембранних цистерн з рибосомами на зовнішній поверхні. Мінімальна товщина цистерн становить 20 нм, але зустрічаються такі, що мають товщину у декілька мікрометрів. В деяких ділянках цитоплазми сплощені цистерни ГрЕПР щільно прилягають одна до одної, утворюючи стопки. Такі ділянки цитоплазми мають назву ергастоплазма (Рис. 7). Комплекс мембран ГрЕПР зв’язаний із зовнішньою мембраною ядерної оболонки і перинуклеарним простором.

Рис. 7. Ергастоплазма
1 - цистерни ГрЕПР з рибосомами, 2 - ядро, 3 - ядерна оболонка, 4 - мітохондрія.
На рибосомах ГрЕПР синтезуються структурні білки, ферменти лізосом і експортні білки. В цистернах ГрЕПР відбувається їхня сегрегація та дозрівання. Тут же відбувається синтез глікопротеїдів і ліпопротеїдів. Інколи ГрЕПР здатний утворювати секреторні гранули, минаючи комплекс Гольджі. Дозрілі білки транспортуються по цистернах ГрЕПР до місця використання. ГрЕПР може утворювати готові мембрани і здійснювати їх вбудовування замість пошкоджених.
Гладкий ендоплазматичний ретикулум (ГлЕПР) - це система анастомозуючих мембранних цистерн, пухирців і трубочок. Зазвичай вони ширші, ніж цистерни ГрЕПР (50-100 нм). Функції ГлЕПР досить різноманітні, це: синтез тригліцеридів (епітелій кишечнику), синтез стероїдних гормонів (коркова речовина надниркових залоз), синтез глікогену, детоксикація (гепатоцити), депо іонів Са2+ (міофібрили) .
Комплекс Гольджі (К. Гольджі, 1898) досить часто розташовується біля ядра поблизу клітинного центра. Являє собою систему диктиосом.
Кожна диктиосома утворена стопкою з 3-10 сплощених і дещо вигнутих цистерн з розширеними кінцями (Рис. 8). В складі диктиосоми цистерни утворюють три основних компартменти: цис-сторона, транс-сторона, проміжний компартмент. З диктиосомою тісно пов’язаний і завжди розглядається разом з нею ще один клітинний компартмент - транс-сітка Гольджі.
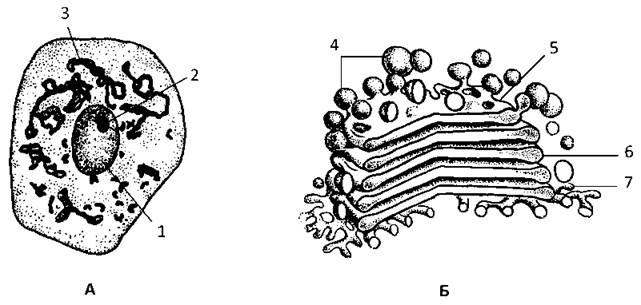
Рис. 8. Будова комплексу Гольджі:
А - нейрони спинного мозку (імпрегнація сріблом за методом Гольджі), Б - ультрамікроскопічнабудова дикитиосоми: 1 - ядро, 2 - ядерце, 3 - комплекс Гольджі, 4 - вакуолі транс-сітки Гольджі, 5 - транс-сторона, 6 - проміжний компартмент, 7 -цис-сторона.
Цис-сторона включає цистерни, звернені до розширених елементів ГрЕПР, а також невеликі транспортні пухирці.
Транс-сторона утворена цистернами, зверненими до вакуолей і секреторних гранул та пухирців.
Проміжний компартмент включає невелику кількість цистерн між цис- і транс-сторонами.
Транс-сітка Гольджі залягає на невеликій відстані від крайової цистерни транс-сторони і бере участь в утворенні лізосом і сегрегації білків для різних транспортних пухирців.
Функції комплексу Гольджі полягають у сегрегації і накопиченні продуктів, синтезованих на ГрЕПР, модифікації секреторного продукту, т.т. його хімічній перебудові та дозріванні, синтезі полісахаридів та мукопротеїдів, упаковці секреторного продукту, утворенні лізосом.
Лізосоми (де Дюв, 1949) - кулястої форми пухирці, розміри та електронна щільність яких коливаються в значних межах. Більшість лізосом все ж таки мають розміри 0,2-0,4 мкм. Матрикс (основна речовина) лізосом містить більше 50 ферментів: рибонуклеази, дезоксирибонуклеази, сульфатази, фосфоліпази, глікозідази, ліпази та ін. Лізосомні ферменти найбільш активні у кислому середовищі (рН ~ 5), для підтримки якого в мембрану лізосоми вбудований протонний насос (Н+,К+-АТФаза). Маркерним ферментом лізосом служить кисла фосфатаза.
Лізосоми беруть участь у внутрішньоклітинному та позаклітинному травленні. Варіанти участі лізосом в перетравлюванні матеріалу внутрішньоклітинних компонентів (аутофагія) або частинок, які потрапили у клітину різними шляхами (гетерофагія), представлені на рис. 9.
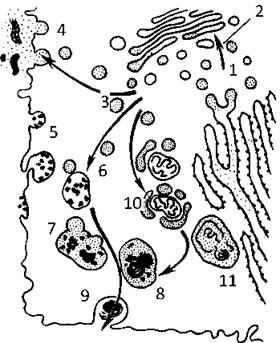
Рис. 9. Участь лізосом у внутрішньоклітинному та позаклітинному травленні:
1 - синтез ферментів лізосом ГрЕПР, 2 - їх переніс до КГ, 3 - утворення первинних лізосом, 4 - позаклітинне травлення, 5 - утворення ендосом, 6 - утворення гетерофагосоми, 7 - вторинна лізосома, 8 - телолізосома (третинна лізосома), 9 - екскреція залишкових тілець, 10 - утворення аутофагосоми, 11 - накопичення ліпофусцину у залишкових тільцях.
У залишкових тільцях досить часто спостерігається накопичення ліпофусцину (пігменту старіння). Це продукт лізосомного перетравлювання, який не піддається подальшому внутрішньоклітинному розщепленню (мембранні пухирці різних розмірів з електроннощільним вмістом, часто містять ліпіди).
Пероксисоми - овальної форми мембранні пухирці розміром 0,11,5 мкм. Матрикс пероксисом зернистий, часто з кристалічною серцевиною. Вони особливо численні в клітинах печінки і нирок. У складі їхньої мембрани містяться специфічні для пероксисом унікальні мембранні білки з М.м. 22 і 70 кД, а в матриксі - більше 40 ферментів, які каталізують анаболічні (біосинтез жовчних кислот) і катаболічні (β-окислення довгих ланцюгів жирних кислот, Н2О2-залежне дихання) процеси. Маркерним ферментом пероксисом служить каталаза. Всі компоненти пероксисом надходять із цитозоля. Тривалість життя пероксисом становить 5-6 діб. Нові пероксисоми виникають із попередніх шляхом їх поділу. Попередні пероксисоми збільшуються у розмірах по мірі надходження до них білків із цитозолю, і від них відокремлюються нові пероксисоми.
Мітохондрії (Бенда, 1898) - органели клітинного дихання, або перетворювачі енергії для внутрішньоклітинних реакцій, займають значну частину цитоплазми (до 20% об.) клітин і концентруються у місцях потреби АТФ: в м’язах - навколо міофібрил, в сперматозоїдах - у футлярі аксонеми.
Припускають, що мітохондрії походять від аеробних симбіонтів, які потрапили в анаеробну еукаріотичну клітину шляхом ендоцитозу і почали брати участь у її окисних процесах. У зв’язку з цим мітохондрії мають власний геном (кільцева ДНК), м-РНК, т-РНК, р-РНК, але більшість мітохондріальних білків кодує ядерна ДНК. Органела функціонує приблизно 10 діб, оновлення мітохондрій відбувається шляхом їх поділу.
Мітохондрії частіше за все мають форму циліндра діаметром 0,1-1 мкм і 7-10 мкм завдовжки. У мітохондрій дві мембрани - зовнішня і внутрішня, яка утворює кристи. Між мембранами знаходиться міжмембранний простір. Внутрішній простір мітохондрії займає матрикс (Рис. 10).
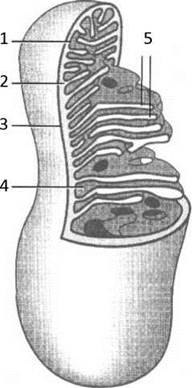
Рис. 10. Мітохондрія:
1 - внутрішня мембрана, 2 — міжмембранний простір, 3 — зовнішня мембрана, 4 - матрикс, 5 - кристи.
Зовнішня мітохондріальна мембрана проникна для багатьох дрібних молекул. Внутрішня проникна вибірково, містить транспортні системи для переносу речовин в обох напрямках і комплекси ланцюга переносу електронів, пов’язані з ферментами окисного фосфорилювання, а також сукцинатдегідрогеназу (СДГ). У міжмембранному просторі накопичуються іони Н+, які викачуються із матриксу при переносі електронів у дихальному ланцюзі, що створює протонний градієнт концентрації по обидва боки внутрішньої мембрани. У матриксі присутні всі ферменти циклу Кребса (крім СДГ), ферменти Р-окислення жирних кислот і деякі ферменти інших систем. В матриксі знаходяться гранули з Mg2+ і Са2+.
Функції мітохондрій:
1. Окислення в циклі Кребса. Матрикс мітохондрії містить ферменти, які окислюють піруват і жирні кислоти до ацетил-КоА, і ферменти, які окислюють ацетил-КоА до СО2. Кінцеві продукти циклу трикарбонових кислот: СО2, що виходить з клітини, і НАДН (нікотинамідаденіндинуклеотид) - джерело електронів, що переносяться дихальним ланцюгом.
2. Транспорт електронів. Електрони транспортуються по дихальному ланцюгу, локалізованому у внутрішній мембрані мітохондрій. Дихальний ланцюг являє собою чотири великих ферментних комплекси (переважно цитохроми) ланцюга транспорту електронів. При проходженні електронів по дихальному ланцюгу іони Н+ відкачуються із матриксу у міжмембранний простір. Таким чином створюється протонний градієнт, енергія якого використовується для синтезу АТФ і транспорту метаболітів і неорганічних іонів у матрикс.
3. Фосфорилювання АДФ. АТФ-синтезуючий комплекс міститься у кристах. При зворотному токові протонів у матрикс через канал у АТФ-синтетазі відбувається фосфорилювання АДФ до АТФ.
4. Спряження окислення і фосфорилювання.Внаслідок спряження цих процесів енергія, що звільняється під час окислення субстратів у циклі Кребса, зберігається у макроергічних зв’язках АТФ. Енергія, запасена у АТФ, реалізується у багатьох процесах функціонування клітини (іонні насоси, рухливість і т.п.). Слід зазначити, що ефективність окисного фосфорилювання у мітохондріях значно вище такої при гліколізі у цитозолі (з одної молекули глюкози під час гліколізу утворюється лише 2 молекули АТФ, у мітохондріях - 36).
5. Розспряження окисного фосфорилювання. Спряження між переносом електронів і фосфорилюванням порушують агенти, які руйнують градієнт Н+ між цитозолем і між мембранним простором або дозволяють Н+ вільно проходити через внутрішню мембрану у матрикс, серед них: 2,4-динітофенол, антикоагулянт дикумарол, жовчний пігмент білірубін. Розспряження фосфорилювання і дихання буває біологічно корисним як спосіб утворення тепла у зимосплячих тварин під час просинання.
6. Теплопродукція. Мітохондрії бурого жиру під час просинання у зимосплячих тварин захоплюють воду, їхній матрикс ущільнюється, міжмембранні простори збільшуються, зовнішні мембрани досить часто лопають, при цьому вони припиняють утворення АТФ, вся енергія окислення субстратів виділяється у вигляді тепла. Ці процеси забезпечує спеціальний розспрягаючий білок. Експресія цього білка стимулюється підвищенням концентрації тироксину, цитозольного кальцію і особливо медіатором симпатичної нервової системи - норадреналіном.
Рибосоми утворюють у клітині дві популяції: цитоплазматичні рибосоми, які містяться у цитозолі, і мітохондріальні рибосоми, які відрізняються від перших своїм складом, розмірами і кількістю (останніх значно менше).
Цитоплазматичні рибосоми або вільно плавають у цитоплазмі (вільні рибосоми) або розташовуються на зовнішній поверхні ГрЕПР (асоційовані рибосоми). Слід зазначити, що саме існування рибосом безпосередньо пов’язане з їхньою функцією, т.т. в клітині не існує непрацюючих рибосом. Як правило, рибосоми об’єднуються при цьому у полірибосоми, або просто полісоми.
У зібраному стані цитоплазматична рибосома має константу седиментації 80S і складається із двох субодиниць (Рис. 11). Мала субодиниця (40S) містить один довгий ланцюг рРНК (близько 2000 нуклеотидів, 18S), з яким зв’язані приблизно 30 рибосомних білків.
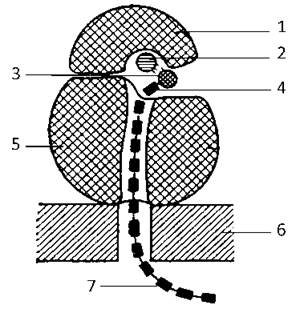
Рис. 11. Схема будови асоційованої рибосоми:
1 - мала субодиниця, 2 - м-РНК, 3 - аміноацил-тРНК, 4 - амінокислота, 5 - велика субодиниця, 6 — мембрана ГрЕПР, 7 - синтезований поліпептидний ланцюг.
Велика субодиниця (60S) містить вдвічі довшу рРНК (4000 нуклеотидів, 28S), з якою зв’язані 2 коротких ланцюги РНК (5,8S і 5S) і близько 45 молекул білків.
Всі рРНК синтезуються у ядерцях, де вони об’єднуються з білками, внаслідок чого утворюються субодиниці рибосом, які являють собою згорнутий рибонуклеопротеїдний тяж, що має декілька функціональних центрів. Зборка рибосом із субодиниць відбувається у цитоплазмі. У непрацюючому стані субодиниці рибосом так і залишаються дисоційованими одна від одної.
Функція рибосом полягає у забезпеченні трансляції - почергового включення амінокислот у поліпептидний ланцюг, що будується у відповідності з послідовністю кодонів у матричній РНК. Насамперед з певним центром малої субодиниці зв’язується початкова ділянка м-РНК. Потім до цього комплексу приєднується тРНК, навантажена першою амінокислотою майбутнього пептидного ланцюга. І лише після цього побудова рибосоми завершується приєднанням великої субодиниці.
Мікротрубочки являють собою нерозгалужені порожнисті циліндри із зовнішнім діаметром у 24 і внутрішнім у 15 нм. Стінка мікротрубочки складається із глобул білка тубуліна, розташованих по пологій спіралі з частотою 13 глобул на 1 виток (Рис 12). Молекула тубуліна - димер із субодиниць α- і β-тубуліна.
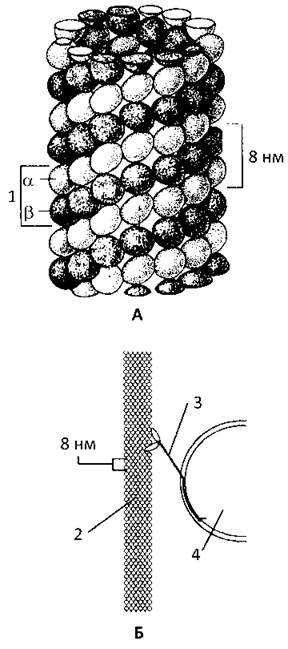
Рис. 12. Будова і функції мікротрубочок.
А. Стінку мікротрубочки утворюють розташовані в один шар димери тубуліну. Вони складаються з двох субодиниць і мають довжину 8 нм.
Б. Білок кінезин перетворює енергію АТФ у механічну роботу. Він забезпечує транспорт органел з одної частини клітини в іншу вздовж мікротрубочок. Крок пересування кінезину становить 8 нм.
1- молекула тубуліну, 2 - мікротрубочка, 3 - кінезин, 4 - органела.
Мікротрубочки полярні. Зборка мікротрубочок здійснюється на плюс-кінці, прикріпленому до якоїсь клітинної структури, шляхом полімеризації тубуліну. Деполімерізація тубуліну на мінус-кінці мікротрубочки приводить до її укорочення. Слід зазначити, що тубулін не має АТФазної активності, тому мікротрубочки до скорочення не здатні.
У клітинах, що не поділяються, мікротрубочки підтримують форму клітини, виконуючи функцію цитоскелету. Мікротрубочки також значною мірою забезпечують транспорт органел і речовин в цитоплазмі клітин, при цьому транспорт здійснюється не всередині, а по поверхні мікротрубочок. Під час мітозу мікротрубочки утворюють веретено поділу, що сприяє правильному розходженню хромосом до полюсів клітини. Крім того, мікротрубочки беруть участь у побудові цілого ряду клітинних структур, а саме: центріолей та аксонем війок і джгутиків.
Важливу роль у зборці та функціонуванні мікротрубочок відіграє т-білок. Зв’язуючись з тубуліном, він стимулює зборку мікротрубочок, а також утворює між ними поперечні зшивки.
Центріолі (Бувре, 1895) - щільні тільця завдовжки 0,3-0,5 мкм з діаметром 0,15 мкм, які можна побачити на межі роздільної здатності мікроскопа.
Центріолі складються із мікротрубочок. Склад кожної центріолі відбивається формулою: (9x3)+0 (Рис. 13). Це означає, що стінка циліндра центріолі складається із 9 периферичних триплетів мікротрубочок, які зв’язані поперечними білковими містками (ручками), а в центрі циліндра мікротрубочки відсутні. Кожний триплет складається із 3-х мікротрубочок (А, В, С). Один оборот спіралі мікротрубочки А містить 13 глобул тубуліну, В і С - по 11. Мікротрубочка А кожного триплету несе дві білкових ручки: внутрішню і зовнішню. Внутрішня ручка прикріплює триплет до розташованої в центрі центріолі білкової втулки діаметром 25 нм, а зовнішня до С- мікротрубочки сусіднього триплету.

Рис. 13. Будова центріолей:
А - електронна мікрофотографія центріолей фібробласта: 1 -центріолі, 2 - світла зона. Б -триплет мікротрубочок: А, В, С - мікротрубочки, 3 - зовнішня ручка, 4 - внутрішня ручка, 5 - втулка.
Центріолі завжди розташовуються парою - під прямим кутом одна до одної. Така структура називається диплосомою, при чому в її складі розрізняють материнську та дочірню центріолі. Навколо диплосоми розташовується зона більш світлої цитоплазми. В ній розташовані сателіти. Від сателітів радіально, як промені, розходяться численні мікротрубочки, через що ця зона називається центросферою. Диплосома разом з центросферою утворюють клітинний центр.
Під час підготовки клітини до поділу утворюються нові центріолі. Це відбувається шляхом дуплікації: кожна із центріолей диплосоми добудовує собі дочірню, при цьому материнська центріоль вихідної диплосоми зберігає за собою і центросферу.
Війки та джутики являють собою цитоплазматичні вирости діаметром близько 0,2 мкм. Вони є спеціальними органелами руху. Висота війок, як правило, не перевищує 5-10 мкм, джгутики ж значно довші - 50-150 мкм.
Скелет або основу війок і джгутиків становить осьова нитка - аксонема. Вона проходить по осі війки або джгутика і утворена із мікротрубочок за формулою: (9x2)+2. Кожний із 9-ти периферичних дуплетів включає 2 мікротрубочки: А і В. Мікротрубочка А має 11 мікротрубочок на виток спіралі, а мікротрубочка В - 13. Від мікротрубочки А в напрямку мікротрубочки В сусіднього дуплету тягнуться 2 ручки. Вони складаються із білка динеїна, що має АТФазну активність. Дві окремі центральні мікротрубочки заключені у центральний футляр. До останнього за допомогою радіальних спиць приєднуються всі дев’ять мікротрубочок А.
При гідролізі АТФ динеїн змінює свою конформацію, при цьому сусідні дуплети дещо переміщуються відносно один одного, що приводить до руху війки, що називається биттям.
В основі аксонеми війок лежить структура, яка теж складається із мікротрубочок - базальне тіло. За будовою базальне тіло фактично являє собою центріоль. Мікротрубочки А і В базального тіла переходять у дуплет аксонеми. Базальним тілом для аксонеми джгутика сперматозоїда служить одна з його центріолей.
Інші фібрилярні структури цитоплазми представлені мікрофіліментами і проміжними філаментами (стара назва - мікрофібрили).
Мікрофіламенти беруть участь у побудові цитоскелету. При цьому вони утворюють у клітині досить густу сітку. Особливо вираженою є кортикальна сітка, яка розташована під плазмолемою. Вона запобігає різкій деформації клітин.
Мікрофіламенти мають товщину 5-7 нм і являють собою подвійну спіраль із глобул білка актину. Філаменти кортикальної сітки за допомогою білка а-актиніну зв’язуюються з білками плазмолеми, білок філамін зв’язує їх між собою.
Утворюючи переважно примембранний цитоскелет, мікрофіламенти забезпечують і деякі рухи клітини: міграція клітин в ембріогенезі, утворення псевдоподій, піно- і фагоцитоз, цитотомія і т.д.
Проміжні філаменти за своїм діаметром (10 нм) займають проміжний стан між мікрофіламентами і мікротрубочками. Білковий склад проміжних філаментів є тканиноспецифічним: в епітеліях вони утворені білком кератином, у клітинах сполучних тканин, ендотелії і гладких міоцитах судин - віментином, у м’язовій тканині - десміном.
Основна функція всіх проміжних філаментів - опорна.
Включення - необов’язкові компоненти цитоплазми, які виникають або зникають залежно від метаболічного стану клітини. Розрізняють включення трофічні (жирові, білкові, вуглеводні), секреторні, екскреторні і пігментні (ендогенні і екзогенні). Секреторні та екскреторні включення, як правило, являють собою мембранні структури. Крім поживних речовин клітина може запасати і мінеральні (залізо у вигляді гемосидерину), в такому випадку говорять про резервні включення.