ЗАГАЛЬНА ГІСТОЛОГІЯ З ОСНОВАМИ ЕМБРІОЛОГІЇ - Долгов О. М. - 2015
Частина І
БІОЛОГІЯ КЛІТИНИ
ЯДРО
Ядро клітини - система генетичної детермінації і регуляції білкового синтезу.
Як відомо, більшість функцій клітини забезпечується білками. Специфічність кожного білка визначається структурою нуклеїнових кислот, які зумовлюють стабільність відтворення структури окремих білків і, в решті решт, всієї клітини в цілому. Ця система генетичної детермінації процесів синтезу білків локалізується саме у клітинному ядрі.
За великим рахунком, ядро забезпечує дві групи загальних функцій: одна - це зберігання і передача генетичної інформації, закодованої триплетним кодом послідовності нуклеотидів ДНК, інша - реалізація цієї інформації, тобто забезпечення синтезу білка.
Щодо першої групи функцій, слід зазначити, що структура ДНК може змінюватись під впливом найрізноманітніших чинників. Зрозуміло, що це може привести до фатальних наслідків для клітини. Стабільність незмінної структури ДНК пов’язана з наявністю репараційних ферментів, які здатні забезпечити виправлення спонтанних пошкоджень молекул ДНК.
З іншого боку, в ядрі відбувається відтворення, або редуплікація, молекул ДНК, що дозволяє шляхом мітозу дочірнім клітинам отримати абсолютно однакові у якісному і кількісному відношенні набори генетичної інформації.
Іншою групою клітинних процесів, пов’язаних з активністю ядра, є створення власне апарату білкового синтезу і регуляція через нього функцій клітини. Сюди слід віднести не тільки транскрипцію на матрицях ДНК різних матричних (інформаційних) РНК (м- або іРНК), всіх видів транспортних (трансферних) і рибосомних РНК (т- та рРНК), але й процесинг, трансляцію та післятрансляційну модифікацію молекул білків. Додамо, що синтезовані у ядерці рРНК тут же комплексуються з рибосомними білками, наслідком чого є утворення рибонуклеопротеїдів (РНП), з яких утворені субодиниці рибосом.
Таким чином, ядро - це не тільки місце, де зберігається генетичний матеріал, але й місце, де він функціонує і відтворюється. Саме через це порушення будь-якої функції ядра закінчується для клітини летальним виходом.
Зазвичай в інтерфазній клітині міститься одне ядро, але бувають і багатоядерні клітини.
Структурними компонентами ядра клітини є хроматин, ядерце, каріоплазма (нуклеоплазма) і ядерна оболонка (Рис. 14).
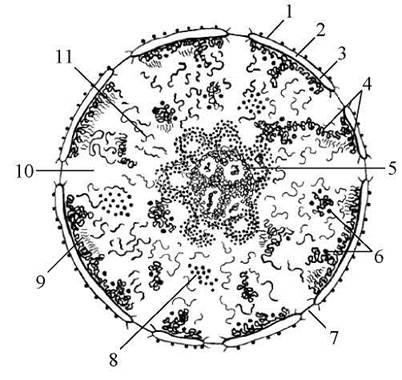
Рис. 14. Схема будови ядра клітини:
1 - зовнішня ядерна мембрана, 2 - перинуклеарний простір, 3 - внутрішня ядерна мембрана, 4 - конденсований хроматин, 5 - ядерце, 6 — перихромати - нові гранули, 7 - комплекс пори, 8 - інтерхроматинові гранули, 9 - перихроматинові фібрили, 10 - каріоплазма, 11 - дифузний хроматин.
Молекула ДНК побудована із двох антипаралельних ланцюгів з комплементарною послідовністю нуклеотидів (Рис. 15). Нуклеотид являє собою фосфатний ефір нуклеозиду. Нуклеотиди за допомогою фосфодиефірних зв’язків утворюють полінуклеотид. Нуклеозиди - N-глікозильні похідні (N-глікозиди) різних азотистих основ, які містять відповідно рибозу або дезоксирибозу. До складу нуклеозидів входять два види азотистих основ: пуринові - аденін (А) та гуанін (G) і піримідинові - цитозин (С), тимін(Т), урацил(и).
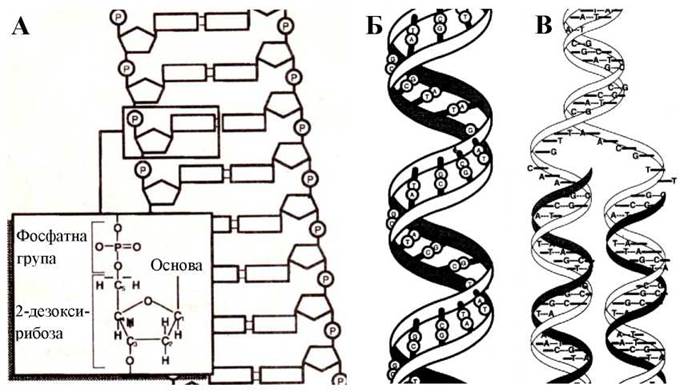
Рис. 15. Структура молекули ДНК.
А. ДНК складається з двох полінуклеотидних ланцюгів, утворених нуклеотидами, з’єднаними фосфодиефірними зв’язками. Ковалентні фосфодиефірні зв’язки з’єднують 5'-атом вуглецю одного нуклеотида з 3'-атомом вуглецю наступного нуклеотида. Б. Антипаралельні ланцюги комплементарно спаровані: аденін (А) з тиміном (Т), гуанін (G) з цитозином (С). В. Перед кожним поділом клітини відтворюють (реплікують) ДНК за допомогою ДНК-полімерази.
Спіраль ДНК (права) являє собою два комплементарних ланцюги полінуклеотидів, з’єднаних водневими зв’язками у парах А-Т і С-G. На один оборот спіралі припадає близько десяти пар азотистих основ.
У складі ДНК виділяють два види послідовностей нуклеотидів - екзони та інтрони. Екзон - кодуюча послідовність нуклеотидів, яка визначає послідовність амінокислот в молекулі білка. Інтрон - некодуюча послідовність між екзонами. Після синтезу РНК на матриці ДНК послідовності РНК, комплементарні послідовностям інтронів, видаляються за допомогою спеціальних ферментних комплексів, а послідовності, що залишились, зшиваються. Виконує цю складну роботу молекулярна «машина», яка має назву сплайсосома (англ. splice - з’єднати), а сам процес визначається терміном сплайсинг.
Кодоном називають послідовність з трьох суміжних нуклеотидів, яка кодує будь-яку амінокислоту або термінацію поліпептидного ланцюга.
Сукупність всіх молекул ДНК клітини прийнято називати геномом, при цьому вважається, що ген - це ділянка молекули ДНК, що кодує завершену послідовність амінокислот у поліпептидному ланцюгові. Експресія (лат. expressio - вираження, проявлення) гена відбувається за схемою: транскрипція (синтез первинного транскрипта на матриці ДНК) → процесинг (утворення мРНК) →трансляція (зчитування інформації з мРНК) → зборка поліпептидного ланцюга (включення амінокислот до поліпептидного ланцюга на рибосомі) → післятрансляційна модифікація (додавання до поліпептиду різних хімічних груп, наприклад, фосфорилювання, карбоксилювання, гідроксилювання тощо) (Рис. 16).

Рис. 16. Схема передачі генетичної інформації (центральна догма молекулярної біології):
А - транскрипція ДНК, Б - сплайсинг, В - транспорт мРНК, Г - трансляція мРНК.
1 - ДНК, 2 - мяРНП, 3 - рибосоми, 4 - білок.
Молекула РНК - полінуклеотид, схожий за хімічним складом з ДНК, але містить у нуклеотидах рибозу замість дезоксирибози та азотисту основу урацил замість тиміну. Розрізняють три форми РНК: мРНК, тРНК, рРНК.
Матрична (інформаційна) РНК (мРНК) складається із сотень і тисяч нуклеотидів. Роль мРНК полягає у переносі генетичної інформації з ядра у цитоплазму і безпосередньої участі у зборці поліпептиду на рибосомі.
Транскрипція мРНК відбувається за допомогою ферменту РНК- полімерази ІІ. Після приєднання до промотора - специфічного сайту молекули ДНК, з якого починається синтез полімеру, РНК-полімераза ІІ розкручує ділянку подвійної спіралі ДНК, оголюючи матрицю для комплементарного спарювання основ. Коли РНК-полімераза зустрічає сигнал термінації транскрипції, синтез полімеру припиняється. Фактично на цьому етапі з ДНК знята РНК-копія, але вона ще не готова до участі у синтезі білка. Поки що це первинний транскрипт, який називають пре-мРНК. У подальшому здійснюється його процесинг (сплайсинг), наслідком якого є утворення мРНК, яка виходить із ядра у цитоплазму.
Як сплайсосома упізнає ділянку пре-мРНК, що підлягає видаленню? На нього вказують спеціальні мітки - сигнальні послідовності нуклеотидів на початку інтронів. Більшість інтронів починаються з пари GU (гуанін-урацил), а закінчуються парою AG (аденін-гуанін). Особливе значення має точка розгалуження - основа А (аденін), яке знаходиться на відстані 20-30 нуклеотидів від кінця інтрону.
Протягом сплайсингу конфігурація сплайсосоми змінюється залежно від стадії процесу. Основними діючими компонентами сплайсосоми є малі ядерні рибонуклеопротеїни (мяРНП), що позначаються як U1, U2, U4, U5 та U6. Вони послідовно з’єднуються з пре-мРНК. Спочатку U1 знаходить місце, де необхідно зробити перший розріз, після цього до так званої точки розгалуження приєднується U2. Утворюється комплекс А. Після приєднання U4, U5 та U6 утворюється комплекс В. Виконавши свої функції, U1 та U4 виходять з комплексу, а сплайсосома переходить в активований стан. На цьому етапі відбувається розрив між інтроном та екзоном, вільний кінець інтрону приєднується до точки розгалуження, і утворюється петля. Активована сплайсосома трансформується у комплекс С, який забезпечує наступний крок - відрізання петлі від другого екзону та зшивання двох екзонів. На останньому етапі вивільняється зріла мРНК, готова для транспортування у рибосому. Малі ядерні рибонуклеопротеїни від’єднуються від вирізаного інтрону і беруть участь у новому циклі.
Під час альтернативного (диференційного) сплайсингу видаляються не тільки інтрони, але й деякі екзони. Результатом є утворення іншого варіанту мРНК, на матриці якої на рибосомі синтезується інший варіант білка. Так, наприклад, у людини, ген структурного білка тропоміозину дає початок п’яти різним варіантам цього білка, які синтезуються у п’яти різних тканинах організму: скелетних м’язах, гладких м’язах, фібробластах, печінці і мозку.
Транскрипцію рРНК і тРНК каталізують відповідно РНК- полімераза І і ІІІ.
Зборку поліпептидного ланцюга під час трансляції запускає стартовий кодон AUG, а припиняють кодони термінації UAA, UAG і UGA.
Транспортна (трансферна) РНК (тРНК) містить близько 80 нуклеотидів. Її роль полягає у доставці амінокислоти до рибосоми, де вони приєднуються до поліпептидного ланцюга, що синтезується. Існує, як мінімум, одна тРНК для кожної з амінокислот. Один кінець тРНК (акцептор) приєднується до амінокислоти, у той час як інший містить антикодон з трьох нуклеотидів, який розпізнає відповідний кодон мРНК і спаровується з ним. Таким чином тРНК перекладає (англ. to translate - перекладати) або трансформує (звідси - трансферна) послідовність нуклеотидів у послідовність амінокислот.
Рибосомна РНК (рРНК) не тільки утворює рибосому в комплексі з білками, вона під час зборки поліпептидного ланцюга взаємодіє з мРНК і тРНК.
Хроматин
Спостереження живих клітин, особливо рослинних, або клітин на фіксованих забарвлених препаратах показали, що усередині ядра виявляються зони щільної речовини, які добре забарвлюються різними барвниками, особливо лужними. Саме ця властивість цього компоненту ядра дозволила Флемінгу (1880) запровадити термін «хроматин» (гр. chroma - барвник, колір). Здатність хроматину сприймати лужні барвники свідчить про його кислотні властивості, які визначаються тим, що до складу хроматину входить ДНК у комплексі з білком. Такі ж самі властивості притаманні і хромосомам, які можна спостерігати під час мітотичного поділу клітин. Таким чином, хроматин у клітинах, що не поділяються, являє собою хромосоми, які втрачають свою компактну форму і деконденсуються.
У клітинах, які не поділяються (інтерфазних), хроматин, що виявляється за допомогою світлового мікроскопа, може рівномірно заповнювати обсяг ядра або ж розташовуватися окремими згустками (хромоцентри). Нерідко він особливо чітко виявляється на периферії ядра (пристінковий, маргінальний, примембранний хроматин) або утворює усередині ядра переплетення досить товстих (близько 0,3 мкм) і довгих тяжів у вигляді внутрішньоядерної мережі (Рис. 17).

Рис. 17. Структурні типи ядер.
А - дифузний, Б - хромоцентричний, В - хромонемний, Г - хромонемно- хромоцентричний, Д - хромосомний, Е - ядро з політенними хромосомами.
Ступінь деконденсації хромосом у ядрах різних клітин може бути різною, і саме вона визначає форми, якими хроматин може бути представленим. Зони, де хромосоми або їхні ділянки виглядають повністю деконденсованими, називають дифузним хроматином, або еухроматином. Зони з неповним розпушенням хромосом називають конденсованим хроматином, або гетерохроматином.
Численними роботами показано, що ступінь деконденсації хроматину в інтерфазі може відбивати функціональне навантаження цієї структури. Чим більш дифузним є хроматин інтерфазного ядра, тим вище в ньому рівень синтетичних процесів. Так, у клітинах лімфоцитів хроматин утворює значні скупчення по периферії клітинного ядра. При стимуляції цих клітин до синтезу ДНК у міру включення попередника ДНК 3Н-тимідину відбувається поступова деконденсація хроматину. У такий же спосіб змінюється структура хроматину при синтезі РНК. Падіння синтезу ДНК і РНК у клітинах звичайно супроводжується збільшенням зон конденсованого хроматину. В еритроцитах нижчих хребетних практично весь хроматин ядер знаходиться в конденсованому стані, і в цих ядрах не відбувається синтезу ні РНК, ні ДНК. Якщо ж ядра цих клітин стимулювати до синтезу РНК, то вони переходять у дифузний стан.
Максимально конденсується хроматин під час мітотичного поділу клітин, коли він виявляється у вигляді тілець - хромосом. У цей період хромосоми не несуть ніяких синтетичних навантажень, у них не відбувається включення попередників ДНК і РНК. Виходячи з цього, можна вважати, що хромосоми клітин можуть знаходитися в двох структурно-функціональних станах: у робочому, частково або цілком деконденсованому, коли з їхньою участю в інтерфазному ядрі відбуваються процеси транскрипції і редуплікації (еухроматин), і в неактивному - у стані метаболічного спокою при максимальній їхній конденсації, коли вони виконують функцію поділу і переносу генетичного матеріалу в дочірні клітини (гетерохроматин). Але в соматичних клітинах зустрічаються ділянки хроматину, які знаходяться у конденсованому стані протягом всього клітинного циклу. У складі таких ділянок розрізняють структурний (конститутивний) і факультативний гетерохроматин. Структурний гетерохроматин міститься в обох гомологічних хромосомах і тому є генетично неактивним, його ще називають облігатним (абсолютним). Розташовується він зазвичай поблизу ядерної оболонки і забезпечує певне просторове розташування генетичного матеріалу в ядрі клітини.
Факультативний гетерохроматин присутній лише в одній з гомологічних хромосом, тому генетична інформація, що міститься в іншій, може бути експресованою. Типовим прикладом факультативного гетерохроматину служить ділянка гетерохроматину, що виявляється в усіх соматичних клітинах генетично жіночого організму ссавців. У 1949 р. Мюррей Барр помітив у ядрах нейронів жінок скупчення хроматину, відсутнє у чоловіків. Коли подібні ділянки гетерохроматину були виявлені практично в усіх соматичних клітинах генетично жіночих організмів, їх стали називати тільцями Барра, або статевим хроматином. До того ж стало відомо, що тільце Барра являє собою одну з Х-хромосом, яка знаходиться у неактивному стані. Процес інактивації Х-хромосоми відомий як лайонизація. Механізм компенсації дози генів Х-хромосоми у жінок пояснює гіпотеза запропонована Мері Лайон. Згідно з нею інактивація Х-хромосоми відбувається в ранньому ембріогенезі випадковим чином (інактивованою може бути або батьківська, або материнська Х-хромосома), захоплює всю Х-хромосому в цілому і характеризується стабільністю, передаючись клітинним нащадкам.
Спостреження за структурою хроматину показали, що його складають елементарні фібрили хроматину (хромонеми) товщиною 25 нм. У хімічному відношенні фібрили хроматину являють собою складні комплекси дезоксирибонуклеопротеїдів (ДНП), до складу яких входить ДНК та спеціальні хромосомні білки у співвідношенні 1:1,3. В складі хроматину виявляється також РНК, кількість якої залежить від синтетичної активності хроматину: в інактивованих ядрах вона виявляється у слідових кількостях, а в активних її кількість в ряду ДНК:білки:РНК може досягати 2.
Довжина індивідуальних лінійних молекул ДНК може сягати кількох сотень мікрометрів і навіть сантиметрів. Так, найбільша перша хромосома людини містить ДНК загальною довжиною у 7 см. Сумарна довжина молекул ДНК однієї соматичної клітини людини становить 170 см при масі всього у 6 · 10-12 г.
Білки хроматину складають 60-70 % його сухої маси. Серед них більше 80 % - гістонові білки. Гістони - лужні білки, збагачені основними амінокислотами, переважно лізином і аргініном. Існує п’ять видів гістонових білків: Н1, Н2а, H2b, H3, H4. Слід відмітити, що гістон Н1 ніколи не виявляється в складі еухроматину. Роль гістонів полягає не тільки у забезпеченні специфічної укладки ДНК у хромосомі, вони беруть участь і в регуляції транскрипції. Гістони розташовуються вздовж молекули ДНК не рівномірно, а блоками. Кожний такий блок - нуклеосому - утворюють вісім молекул гістонів, навколо яких у два оберти упакована частина молекули ДНК (Рис. 18).

Рис. 18. Нуклеосома в еухроматині.
Нуклеосома вміщує по дві копії гістонових білків Н2а, Н2b, H3, H4. У конденсованому хроматині нуклеосоми з’єднані гістоном Н1.
Поперечний розмір нуклеосоми становить близько 10 нм. Під час утворення нуклеосом відбувається компактизація ДНК, за рахунок чого вона коротшає у 5 разів. Хромосомна фібрила, що утворюється внаслідок такої компактизації, нагадує нитку бус або чоток, де кожна бусина є нуклеосомою. Нуклеосоми розділені інтервалами у 200 пар азотистих основ. Такі хромосомні фібрили додатково поздовжньо конденсуються, утворюючи основну елементарну фібрилу хроматину завтовшки 25 нм (Рис. 19).
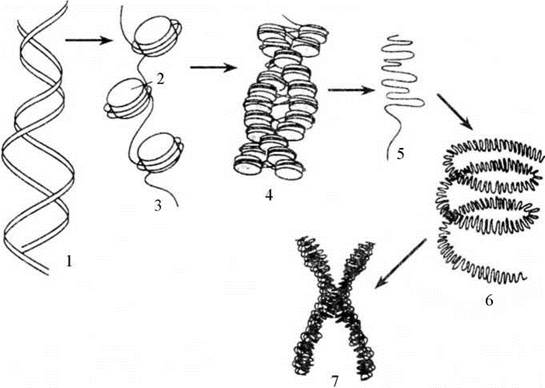
Рис. 19. Організація хроматину.
1 - молекула ДНК, 2 - нуклеосома, 3 - хромосомна фібрила, 4 - елементарна фібрила хроматину, 5 - еухроматин, 6 - гетерохроматин, 7 - метафазна хромосома.