Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Структура РНК
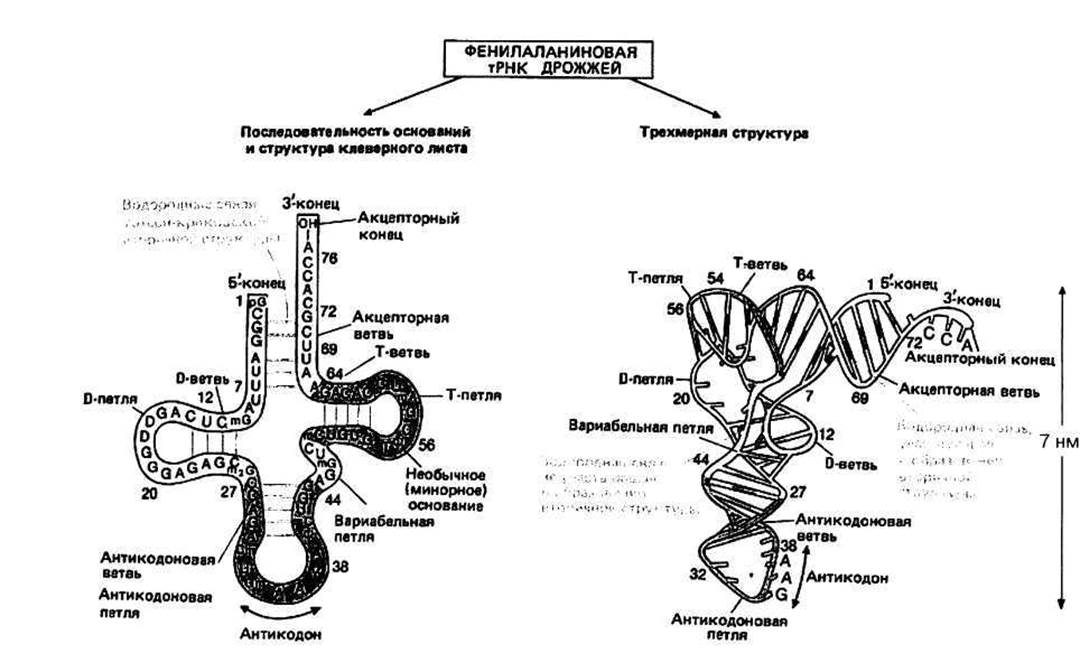
Рис. 19.1.
Рибонуклеиновые кислоты (РНК), присутствующие в клетках как про-, так и эукариот, бывают трех основных типов: информационная (матричная, мРНК), транспортная (тРНК) и рибосомная (рРНК).
В ядре клеток эукариот содержится РНК четвертого типа — гетерогенная ядерная РНК (гяРНК). У некоторых вирусов РНК служит носителем генетической информации (гл. 5). мРНК является копией (транскриптом) соответствующей ДНК. Этот транскрипт служит матрицей для синтеза белка. Каждые три последовательных основания мРНК (называемые кодоном) детерминируют один аминокислотный остаток. Молекулы тРНК переносят специфические аминокислотные остатки к определенному участку мРНК в ходе синтеза белка. В настоящее время структура тРНК хорошо известна (см. ниже). Молекулы рРНК встречаются в различных формах и образуют в комплексе с белками рибосому (гл. 24) — сложную органеллу, в которой происходит синтез белка.
Сведберг (S)-единица измерения коэффициента седиментации, являющегося мерой массы макромолекулы (гл. 44). Его находят, измеряя скорость осаждения молекул в центробежном поле. Значения коэффициента седиментации РНК Е. coli приведены в табл. 19.1.
Транспортная РНК (тРНК) узнает соответствующий кодон в мРНК и переносит нужную аминокислоту к растущей полипептидной цепи. Узнавание кодона в мРНК осуществляется с помощью трех последовательных оснований в тРНК, называемых антикодоном. Аминокислотный остаток может присоединяться к 3'-концу молекулы тРНК. Специфичность такой системы переноса обеспечивается тем, что имеется по крайней мере одна тРНК для каждой аминокислоты. Так, тРНК для Phe обозначается тРНКРЬе.
Таблица 19.1. Характеристики разных РНК Е. coli
|
Тип РНК |
Содержание, % |
Коэффициент седиментации, S |
Мол. масса, Мг |
Число нуклеотидов |
|
рРНК |
80 |
23 |
1 000 000 |
3000 |
|
6 |
500 000 |
1500 |
||
|
5 |
35 000 |
100 |
||
|
тРНК |
15 |
4 |
25 000 |
75 |
|
мРНК |
5 |
4-26 |
25 000-1000 000 |
75-3000 |
Химическая структура тРНК. Молекула тРНК состоит примерно из 75 нуклеотидов, ковалентно связанных друг с другом в линейную цепочку с мол. массой Мг = 25000 и коэффициентом седиментации 4S. Установлены нуклеотидные последовательности многих тРНК; первая из них — для тРHКAlа из дрожжей — была определена Холл и в 1965 г. Все тРНК начинаются с фосфорилированного 5'-конца; первым основанием обычно является G. На 3'-конце всегда присутствуют три основания — ССА и концевая 3'- гидроксильная (—ОН) группа. Составляющими тРНК являются не только четыре обычных основания А, С, G и U, но также несколько необычных, или минорных, оснований. Например, в дрожжевой тРНКРhе минорными основаниями являются псевдоуридин (обозначаемый Ψ), дигидроуридин (D) и основание, обозначаемое Y. Кроме того, в некоторых обычных основаниях один или два атома водорода могут быть замещены на метильные (СН3) группы; метилированные или диметилированные основания отмечаются соответственно с помощью символов m или т2 при букве, обозначающей то или иное основание.
Клеверный лист — это структурное представление молекулы тРНК, полученное из условия образования максимального числа уотсон-криковских пар оснований при данной нуклеотидной последовательности. Участки, в которых с помощью водородных связей образовались такие пары оснований, называются стеблями, а одноцепочечные участки — петлями. Все известные тРНК образуют «клеверный лист» с четырьмя стеблями (акцепторным, D, антикодоновым и Т) и тремя петлями (D, антикодоновой и Т). Некоторые тРНК имеют дополнительные петли и/или стебли (например, вариабельная петля дрожжевой тPHKPhe).
Трехмерная структура дрожжевой тРНКPhe была установлена с помощью рентгеноструктурного анализа кристаллов этой тРНК. Термин «вторичная структура» относится к расположению в пространстве атомов внутри стебля. Третичная структура - это пространственное расположение всех частей молекулы. Вторичная структура Каждый стебель состоит из двух антипараллельных цепей, основания которых образуют друг с другом уотсон-криковские пары с помощью водородных связей. Такое пространственное расположение, установленное экспериментально, было правильно предсказано исходя из структуры клеверного листа. Стебли имеют форму правой двойной спирали, известной как А-форма РНК.
Двойная спираль РНК в А-форме содержит 11 пар оснований на виток, шаг спирали равен 3,1 нм. Расстояние между соседними парами оснований вдоль оси спирали составляет 0,28 нм (3,1 нм/11); они повернуты друг относительно друга на угол 33° (360°/11). Плоскости оснований составляют ~20° с нормалью к оси двойной спирали. А-форма РНК близка к А-форме ДНК (гл. 18). РНК не способна переходить в В-форму. Это обусловлено присутствием в ней объемной 2'- гидроксильной группы рибозы, которой нет в дезоксирибозе.
Третичная структура тРНК. Молекула напоминает по форме букву Г. Она значительно более вытянута, чем глобулярные белки той же молекулярной массы. Акцепторный и Т-стебли уложены в пространстве таким образом, что образуют одну непрерывную спираль — «перекладину» буквы Г; антикодоновый и D-стебли образуют «ножку». Эти две части содержат каждая около 10 пар оснований. Почти все основания в тРНК участвуют в вандерваальсовых и гидрофобных взаимодействиях, стабилизирующих пространственную структуру молекулы.
В тРНК имеются разнообразные водородные связи. В стеблях, являющихся двойными спиралями, образуются уотсон-криковские водородные связи. Многие основания, расположенные вне стеблей, образуют водородные связи с другими основаниями, но эти связи отличаются от уотсон-криковских. Помимо этого 2'-гидроксильные группы полинуклеотидной цепи также образуют водородные связи — с основаниями и с атомами водорода основной цепи.
Антикодон молекулы тРНК - это три последовательных основания, с помощью которых распознается соответствующий кодон мРНК. Узнавание осуществляется путем образования уотсон-криковских водородных связей между основаниями кодона, с одной стороны, и антикодона — с другой, при условии что полинуклеотидные цепи антипараллельны (рис. 19.2).
![]()
Рис. 19.2.
ДРУГИЕ МОЛЕКУЛЫ РНК. К таким молекулам относятся мРНК и одноцепочечная вирусная РНК. Их нуклеотидная последовательность также была определена экспериментально. Эти исследования показали, что отдельные участки молекул РНК могут, подобно тРНК, образовывать шпильки, стабилизированные водородными связями. Двухцепочечная вирусная РНК, по-видимому, находится в А-подобной форме. Для более подробного изучения строения всех этих молекул необходимы рентгеноструктурные исследования.
СЕКВЕНИРОВАНИЕ РНК в настоящее время может осуществляться очень быстро, причем сразу для участков длиной в несколько сотен оснований. Раньше эта процедура была весьма длительной и трудоемкой и можно было определять последовательность участков РНК длиной не более 100 оснований (как в работе Холли 1965 г.; см. выше). Ныне для этой цели используется ряд методов. Здесь применяют два основных подхода — прямой подход и метод копирования.
В случае прямого подхода определяют последовательность оснований в самой РНК. Сначала метят 5'- или 3'-конец молекулы РНК с помощью 32РО4. Затем меченую цепь расщепляют с помощью ферментов, специфичных к определенным последовательностям. Проводя гель-электрофорез и радиоавтографию, получают карту, по которой находят последовательность оснований, подобно тому как это делается для ДНК (гл. 18).
В методе копирования вначале синтезируют цепочку ДНК, комплементарную РНК, и затем с помощью методов, описанных в гл. 18, определяют ее нуклеотидную последовательность. Для получения комплементарной цепи ДНК обычно используют обратную транскриптазу. Это фермент обнаружен в некоторых вирусах и осуществляет обратную транскрипцию, т. е. катализирует процесс синтеза ДНК (обозначаемой кДНК), комплементарной РНК-матрице. (Обратная транскрипция — это процесс, играющий ключевую роль и при клонировании генов; см. гл. 30.) Пусть, например, какой-то участок РНК имеет последовательность 3'- GGGUCA. Тогда кДНК будет иметь последовательность 5'-GCCAGT. Применив метод химического расщепления для секвенирования ДНК (гл. 18), получают радиоавтограмму, представленную на рис. 19.3.

Рис. 19.3.