Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Транскрипция ДНК
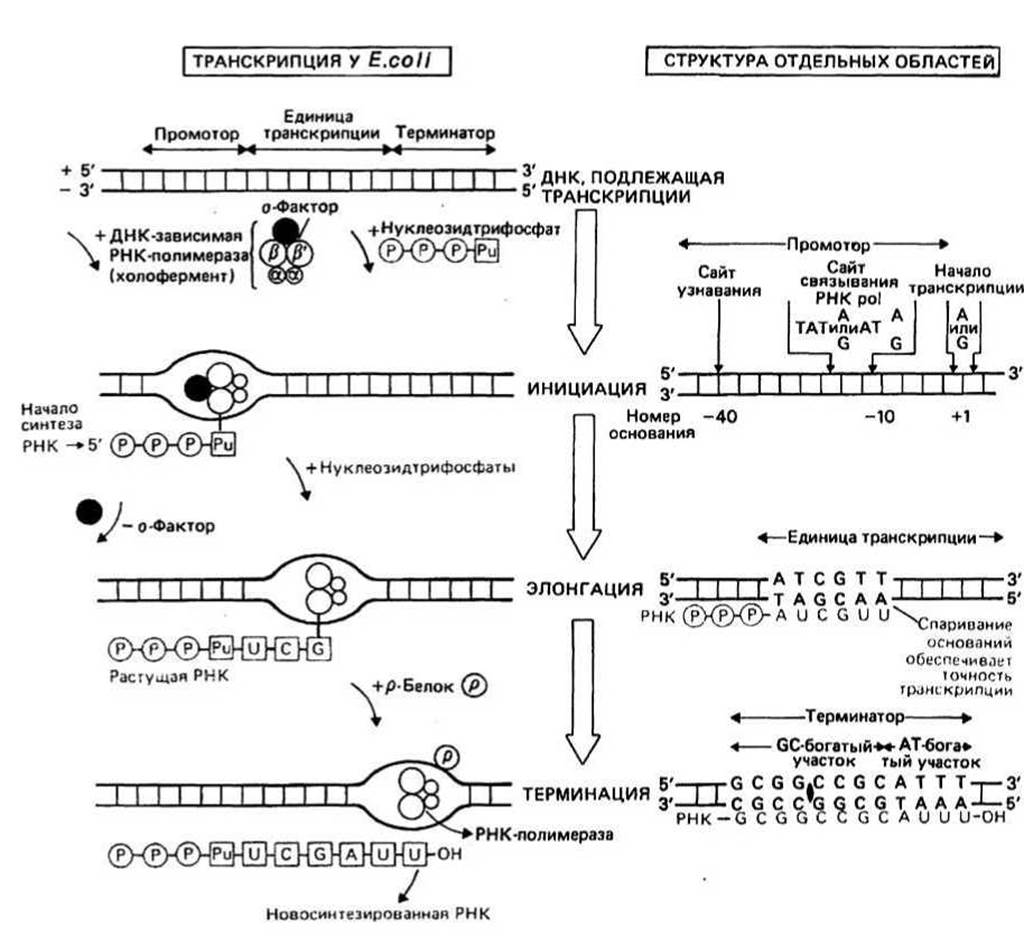
Рис. 22.1.
Транскрипция — процесс переноса генетической информации от ДНК к РНК. Все виды РНК - мРНК, рРНК и тРНК - синтезируются в соответствии с последовательностью оснований в ДНК, служащей матрицей. Транскрибируется только одна, так называемая «+»-цепь ДНК. Процесс транскрипции у прокариот и эукариот существенно различается.
ТРАНСКРИПЦИЯ У ПРОКАРИОТ
ДНК-зависимая РНК-полимераза — это фермент, катализирующий синтез РНК. Он состоит из несколько субъединиц: двух а, одной ß, одной ß' и одной σ. Их комплекс называется холоферментом (a2ßß'σ) и имеет мол. массу (Мг) около 500 000. Фермент, лишенный с-субъединицы, называется корферментом.
Для инициации транскрипции необходимы холофермент, нуклеозидтрифосфат (всегда АТР или GTP) и наличие специального участка в ДНК, называемого промотором. Когда полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и образуется открытый промоторный комплекс.
Промотр - это участок молекулы ДНК, имеющий размер около 40 пар оснований и расположенный непосредственно перед участком инициации транскрипции. Синтез РНК всегда начинается с оснований А или G в «+»-цепи ДНК. Участок связывания холофермента расположен «левее» сайта инициации (в направлении 3' - 5' в «+»-цепи) на расстоянии примерно 10 оснований. Если сравнить последовательности оснований «+»-цепи ДНК у разных промоторов, то мы обнаружим, что они весьма близки, хотя и не идентичны. Эта так называемая последовательность Прибнова имеет вид TATPuATPu, где Рu означает пурин (А или G). Таким образом, холофермент связывается со специфической последовательностью или группой последовательностей. Обычно на расстоянии около 40 оснований «левее» участка инициации находится второе место связывания РНК-полимеразы, где, как полагают, происходит связывание а-субъединицы с ДНК.
Элонгация цепи РНК — это та стадия транскрипции, которая наступает после присоединения примерно восьми рибонуклеотидов. В этот момент РНК-полимераза претерпевает структурное изменение, при котором от комплекса отделяется с-субъединица и остается корфермент (a2ßß'), катализирующий дальнейшее удлинение цепи РНК. При этом к цепи присоединяются те рибонуклеозидтрифосфаты, которые обеспечивают правильное спаривание с «—»-цепью ДНК. Движущийся вдоль ДНК корфермент действует подобно застежке-молнии, «раскрывая» двойную спираль, которая замыкается позади фермента по мере того, как соответствующие основания РНК спариваются с основаниями ДНК в «—»-цепи. «Раскрытая» ферментом область простирается только на несколько пар оснований. На рис. 22.2 представлена химическая реакция удлинения цепи РНК от 3'-конца.
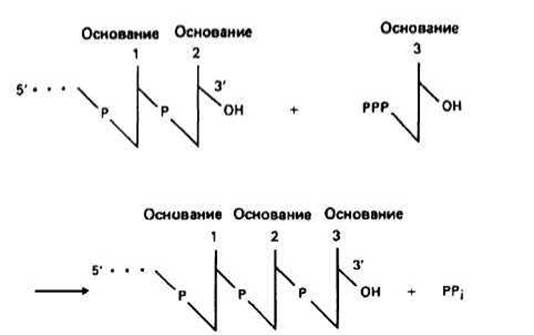
Рис. 22.2.
Терминация (прекращение роста) цепи РНК происходит на специфических участках ДНК, называемых терминаторами. Начало этих участков обычно обогащено GC-парами, а остальная последовательность — АТ-парами. GC-богатый участок часто представляет собой палиндром. Это означает, что при движении вдоль «+»-цепи в одном направлении, а вдоль «—»-цепи - в противоположном читается одна и та же последовательность оснований. В остановке синтеза РНК именно на терминаторе важную роль играет р-белок.
Посттранскрипционный процессинг — это процесс созревания, при котором первичный РНК-транскрипт модифицируется и превращается в зрелую РНК. Характер и степень модификации РНК зависят от типа РНК.
Молекулы мРНК у прокариот не подвергаются процессингу. У некоторых бактерий транскрипция и трансляция сопряжены, т. е. происходят одновременно. 5'-конец мРНК может транслироваться на рибосоме и затем подвергаться деградации еще до завершения синтеза ее 3'-конца. Молекулы тРНК вначале синтезируются в виде про-тРНК, которая примерно на 20% длиннее, чем соответствующая тРНК. Лишние последовательности, расположенные у 5'- и 3'- концов, удаляются с помощью таких ферментов, как рибонуклеазы Q и Р. Иногда молекула про-тРНК состоит из двух или более молекул тРНК, соединенных между собой. Их разделение также осуществляется с помощью рибонуклеаз. Если 3’-конец тРНК не несет концевой последовательности ССА, то эти основания присоединяются при постсинтетической модификации. Все тРНК содержат минорные основания (гл. 19), которые являются химически модифицированными формами четырех главных оснований (А, С, G и U). Эта модификация происходит после завершения транскрипции.
Гены рРНК прокариот расположены в транскрипционных блоках. Три генарРНК Е. coli (16S, 23S и 5S) (гл. 24) располагаются вместе с генами нескольких тРНК в одном таком блоке и транскрибируются в виде одной молекулы РНК. Эти молекулы рРНК и тРНК отделены друг от друга спейсерной РНК. Расщепление первичного транскрипта на отдельные составляющие катализирует рибонуклеаза Q; поскольку этот фермент специфичен к двухцепочечной РНК, предполагают, что в области спейсеров образуются двухцепочечные шпильки, которые фермент узнает и вырезает.
ТРАНСКРИПЦИЯ И ЭУКАРИОТ
У эукариот для транскрипции используются три ДНК - зависимых РНК-полимеразы. Полимераза I локализована в ядрышке, где она катализирует синтез рРНК в виде большого первичного транскрипта, содержащего молекулы рРНК 18S, 5,8S и 28S. Полимераза II находится в нуклеоплазме и, вероятно, участвует в синтезе первичного транскрипта мРНК. Полимераза III также локализована в нуклеоплазме и участвует в синтезе тРНК и 5S-pPHK.
Синтез РНК включает стадии инициации, элонгации и терминации, но в этих процессах часто принимают участие другие ферменты и последовательности оснований, чем у прокариот. Например, промоторные последовательности у эукариот отличаются от таковых у прокариот. Однако первыми основаниями, включаемыми в РНК при инициации, являются, как и у прокариот, А или G.
Молекулы мРНК обычно образуются из больших по размеру молекул-предшественников, называемых гетерогенной ядерной РНК (гяРНК). Для образования зрелой мРНК эти молекулы подвергаются модификации по 5'- и 3'-концами и сплайсингу. После такой модификации транскрипты переносятся из ядра в цитоплазму.
Сплайсинг гяРНК — это удаление последовательностей РНК, соответствующих интронам ДНК, и соединение участков, которые транскрибированы с кодирующих последовательностей (экзонов) (гл. 27). Место сплайсинга должно быть определено с высокой точностью, поскольку ошибка даже в одно основание приведет к синтезу белка с неправильной аминокислотной последовательностью. Такая специфичность сплайсинга обеспечивается строго определенной последовательностью оснований в интроне, отвечающей обычно основаниям GU или GA в начале соответствующей РНК и основаниям AG - в конце.
Модификация 5'-конца мРНК приводит к образованию особой последовательности, называемой кэпструктурой. При модификации 3'-конца к нему присоединяется последовательность poly(A) длиной 150-200 нуклеотидов.
Процессинг тРНК у эукариот протекаетпо такому же механизму, как и у прокариот. Функционально активные молекулы образуются из более длинного предшественника, который подвергается расщеплению и модификации с включением минорных оснований.
Процессинг рРНК также аналогичен соответствующему процессу у прокариот. Первичный транскрипт содержит участки, отвечающие 18S-, 5,8S- и 28S-рРНК (гл. 24), разделенные спейсерами. Как и у прокариот, эти три рРНК образуются при расщеплении спейсерных последовательностей.