Введение в молекулярную биологию: От клеток к атомам - Энтони Рис, Майкл Стернберг 2002
Нуклеиновые кислоты и гены
Регуляция экспрессии генов
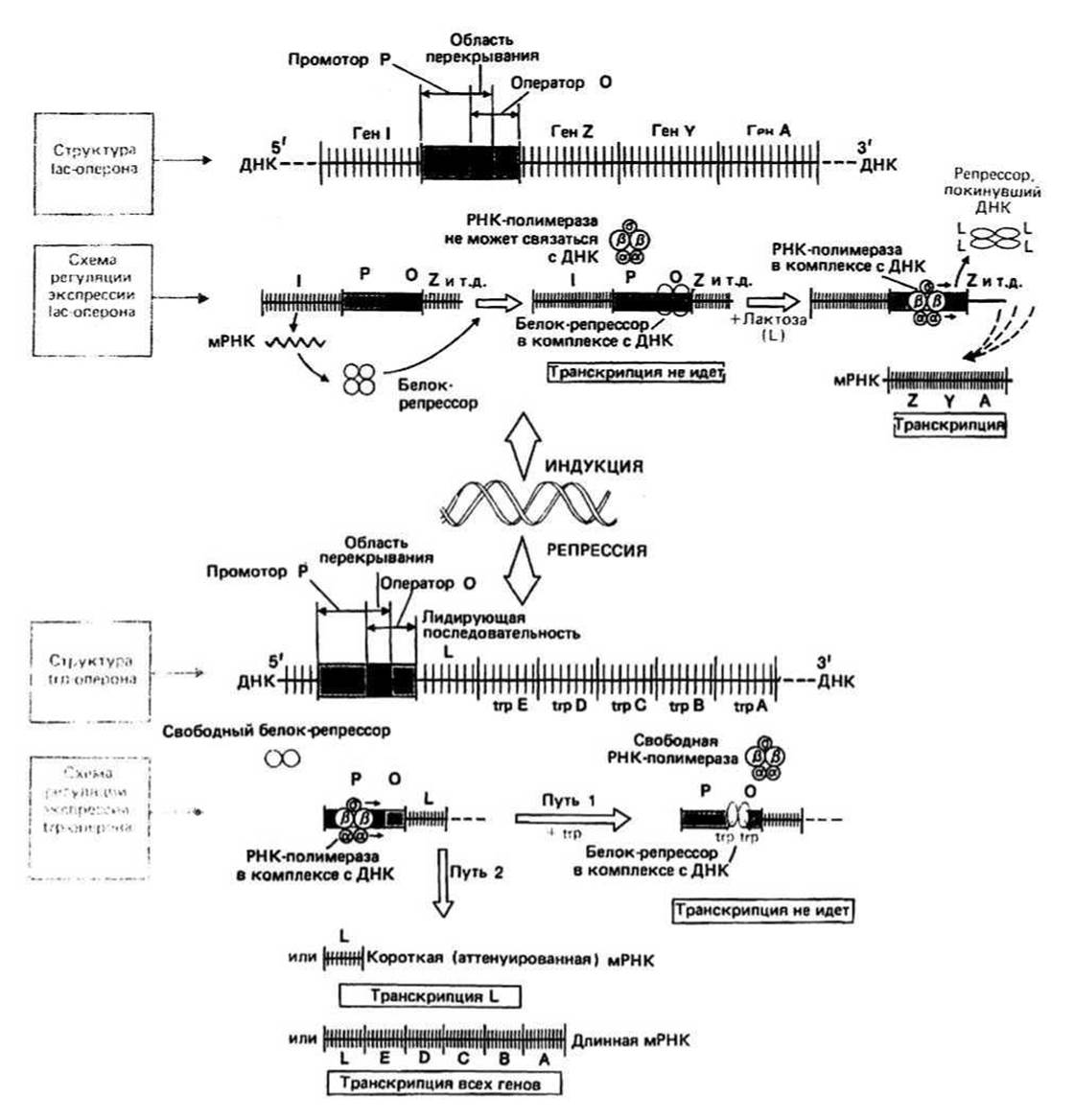
Рис. 28.1.
Экспрессия генов как у прокариот, так и у эукариот регулируется при помощи целого ряда механизмов. Некоторые из механизмов такого рода, действующие в бактериальных системах, изучены довольно хорошо, и два из них будут рассмотрены ниже, но о том, как действуют регуляторные механизмы в клетках эукариот, известно немного.
Прокариоты — это простейшие одноклеточные организмы, которым для того, чтобы выжить, требуется лишь благоприятная химическая среда. Если для обеспечения жизнедеятельности клетке необходим какой-то метаболит, она должна быть способна к синтезу ферментов, которые «пристроят» его в нужное место. Однако синтезировать такие ферменты в отсутствие соответствующего метаболита было бы для клетки расточительством.
Предположение об индукции синтеза ферментов было высказано Жакобом и Моно в 1961 г. для того, чтобы объяснить, каким образом бактериальные клетки реагируют на изменения окружающей их среды. После введения в эту среду лактозы (молочного сахара) в качестве субстрата содержание в бактериальной клетке (ß-галактозидазы — фермента, участвующего в расщеплении лактозы, — увеличивается в 100 раз. Такая активация транскрипции называется индукцией. Одновременно с лактазой индуцируются еще два белка: галактозидпермеаза (белок, осуществляющий транспорт лактозы через мембрану) и тиогалактозид-ацетилтрансфераза. Три структурных гена, кодирующих эти белки, обозначаются соответственно буквами Z, Y и А и вместе с операторным участком образуют так называемый lac-оперон. Было показано, что транскрипционная активность входящих в оперон генов регулируется четвертым, регуляторным геном. Ген-регулятор (ген I) в lac-системе расположен рядом со структурными генами Z, Y и А. Существование гена I было доказано генетическими методами. Когда были выделены мутантные бактерии, лишенные гена I (I- -бактерии), то оказалось, что в таких бактериях экспрессия генов Z, Y и А поддерживается на высоком уровне даже в отсутствие лактозы, т. е. происходит конститутивно. После того как в I- - бактерии ввели фрагмент ДНК, содержащий ген I, экспрессия генов Z, Y и А вновь стала чувствительной к присутствию лактозы. Отсюда был сделан вывод о том, что ген I кодирует какое-то диффундирующее регуляторное вещество, названное репрессором.
Репрессор — это белок, блокирующий транскрипцию гена. В lac-системе репрессор представляет собой тетрамерный белок и называется lac-репрессором. Он связывается с определенным участком на ДНК, который называется оператором.
Оператор(О) представляет собой небольшой участок ДНК, граничащий с первым структурным геном. Белок-репрессор может связываться с этим участком, блокируя тем самым инициацию транскрипции. Операторная последовательность, с которой связывается репрессор, содержит участок палиндромной ДНК. Последовательность с осью симметрии 2-го порядка, изображенная на рис. 28.2, является частью места связывания с репрессором в lac - опероне.
Промотор (Р) - это небольшой участок ДНК перед оператором. Он служит местом связывания РНК-полимеразы. Место связывания репрессора (О) и участок Р слегка перекрываются, так что, когда репрессор находится на ДНК, РНК-полимераза не может связаться с промотором и транскрипция не идет.
Индуктор - представляет собой низкомолекулярное вещество, которое связывается с репрессором и переводит его в неактивную форму, неспособную более связываться с оператором. Так, в /ас-системе индуктором является лактоза, после ассоциации с которой репрессор отсоединяется от /яс-оператора. Индукция является одной из форм негативной регуляции, называемой так потому, что транскрипция может идти лишь после удаления репрессора. Еще одной разновидностью негативной регуляции является так называемая катаболитная репрессия.
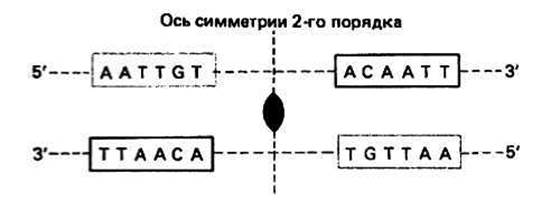
Рис. 28.2.
Репрессия происходит тогда, когда репрессор связывается с оператором не иначе, как в комплексе с низкомолекулярным кофактором (корепрессором). Таким корепрессором часто бывает конечный продукт белкового синтеза, кодируемый опероном. Тогда, если концентрация этого продукта становится слишком высокой, он связывается с репрессором и дальнейший его синтез прекращается. Примером такой системы может служить триптофановый оперон.
Триптофановый оперон состоит из оператора и пяти структурных генов (А— Е). Последние кодируют ферменты, участвующие в биосинтезе триптофана, одной из незаменимых аминокислот. По мере увеличения концентрации триптофана наступает момент, когда дальнейший его синтез становится нежелательным и транскрипция прекращается. «Выключение» транскрипции происходит следующим образом. Триптофан связывается с димерным репрессором (trp-репрессором), который кодируется отдельным, не входящим в оперон регуляторным геном. При этом происходит конформационное изменение, и открывается участок, способный связываться с операторной последовательностью в ДНК. Весь комплекс связывается далее с ДНК и блокирует место связывания с РНК-полимеразой (промотор). Это пример регуляции с помощью репрессии (путь 1 на рис. 28.1).
Вариация длины транскрипта — еще один механизм, посредством которого может осуществляться регуляция экспрессии оперонов в бактериях. Так, в trp - опероне помимо репрессии используется и другая система регуляции. Она связана с наличием в ДНК участка, расположенного непосредственно перед первым структурным геном (trpE) и называемого аттенуатором (путь 2 на рис. 28.1). Аттенуатор представляет собой нуклеотидную последовательность, содержащую команду, по которой происходит преждевременная терминация транскрипции. В случае trp-оперона при высокой концентрации триптофана 90% всех транскриптов терминируется после транскрипции всего лишь 140 нуклеотидов и до начала транскрипции структурных trp-генов.
Позитивная регуляция — еще один способ регуляции экспрессии гена (на рисунке не показан). Он отличается от негативной регуляции тем, что транскрипция «включается», а не «выключается» после присоединения регуляторного белка к оперону. Корепрессор, или скорее коактиватор, присоединяется к белку- активатору; далее весь комплекс связывается с соответствующим участком на ДНК, и лишь после этого может происходить транскрипция. Примером регуляции такого типа может служить процесс, происходящий при участии катаболитного белка-активатора, коактиватором которого является сАМР (см. дополнительную литературу). В отсутствие (или при низкой концентрации) сАМР, что имеет место при размножении бактерий в присутствии глюкозы, блокируется транскрипция некоторых оперонов, в частности lac-оперона. Такое явление называется катаболитной репрессией.
ЭУКАРИОТИЧЕСКИЕ ОРГАНИЗМЫ в значительной части представлены многоклеточными формами с высокой специализацией клеток. Хотя во всех клетках человека содержится совершенно одинаковая ДНК, в разных тканях происходит экспрессия далеко не одинаковых наборов генов. Таким образом, должны существовать какие-то механизмы, при участии которых одни гены работают (т. е. экспрессируются), а другие нет. Для этой цели используется целый ряд механизмов: регуляция на уровне транскрипции и на уровне трансляции, посттранскрипционная и посттрансляционная регуляция и регуляция с помощью гормонов.
Регуляция на уровне транскрипции осуществляется при синтезе мРНК. Средние концентрации индивидуальных мРНК, транскрибируемых с разных генов, сильно отличаются друг от друга. Это обусловлено тем, например, что мРНК-копии одних генов разрушаются быстрее других, либо тем, что их синтез происходит медленнее. Регуляция может осуществляться при помощи белков, способных связываться с ДНК, и даже при помощи коротких фрагментов РНК, которые спариваются с ДНК, предположительно блокируя места прикрепления РНК-полимеразы. Таким образом скорость транскрипции может снижаться или, наоборот, повышаться.
Посттранскрипционная регуляция осуществляется на уровне процессинга мРНК. Даже в том случае, если транскрипция двух разных генов проходит с одинаковой скоростью, дальнейший процессинг мРНК, включающий модификацию 5'- и 3'-концов и сплайсинг экзонов, может протекать по-разному у разных мРНК.
Регуляция на уровне трансляции осуществляется за счет того, что исключается возможность использования мРНК в качестве матрицы для синтеза белка, хотя она и присутствует в цитоплазме. В ооците морского ежа, например, много мРНК, но сколько-нибудь заметного синтеза белка не происходит до тех пор, пока ооцит не будет оплодотворен. Лишь после этого молекулы мРНК подвергаются модификации, т. е. «приобретают» шпильку на 5'-конце (так называемый кэп, или «шапочку») и «шлейф» из poly (А) на 3'-конце (гл. 22), и могут далее включаться в нормальный трансляционный процесс, который завершается построением молекулы белка.
Посттрансляционная регуляция основана на том, что многие белки синтезируются в неактивной форме и должны еще пройти стадию модификации. Так, в р- клетках поджелудочной железы синтезируется не инсулин как таковой, а его предшественник, полипептидная цепь которого длиннее инсулиновой и содержит еще некоторую добавочную последовательность аминокислотных остатков. Лишь после того как эта последовательность вырезается протеолитическим ферментом, получается собственно гормон в своей функциональной форме. Таким образом, производство активного гормона может регулироваться посттрансляционным путем через регуляцию активности протеолитического фермента.
Регуляция с помощью гормонов - это частный случай регуляции на уровне транскрипции; таким путем организм заставляет клетку «включить» определенные гены в ответ на внешний стимул. Так, стероидные гормоны из тех клеток, где они были синтезированы, попадут в конечном счете в цитоплазму соответствующих клеток-мишеней, откуда специальный транспортный белок перенесет их в ядро, где они смогут активировать те или иные гены путем прямого взаимодействия с хроматином в соответствующих местах. Каждый гормон активирует свой набор генов. Таким способом обеспечивается соответствие реакции клетки роду сигнала, который клетка воспринимает через посредничество соответствующего гормона (гл. 41).